

Endothelial pannexin 1-TRPV4 channel signaling lowers pulmonary arterial pressure in mice
- PMID: 34490843
- PMCID: PMC8448527
- DOI: 10.7554/eLife.67777
Endothelial pannexin 1-TRPV4 channel signaling lowers pulmonary arterial pressure in mice
Abstract
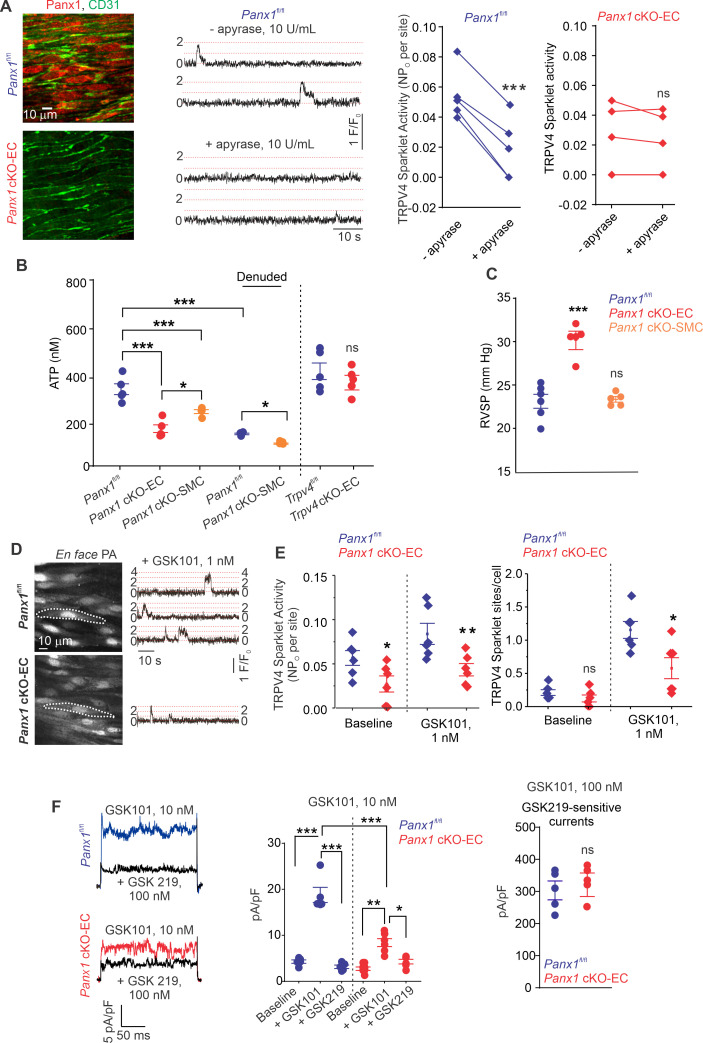
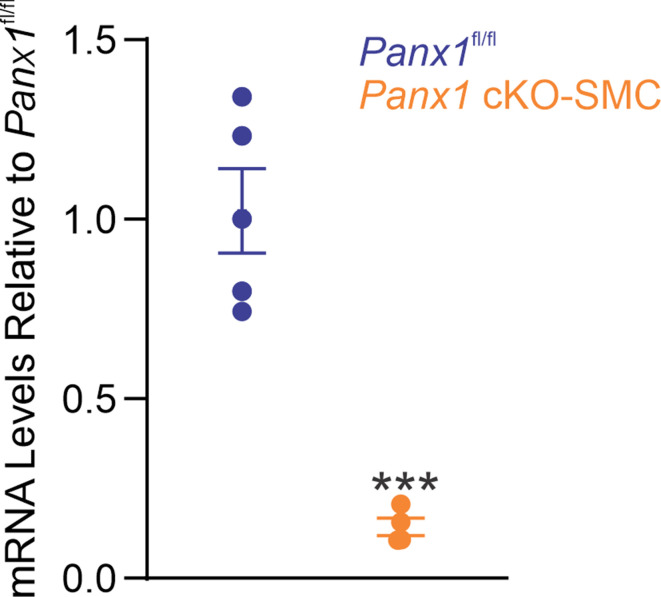
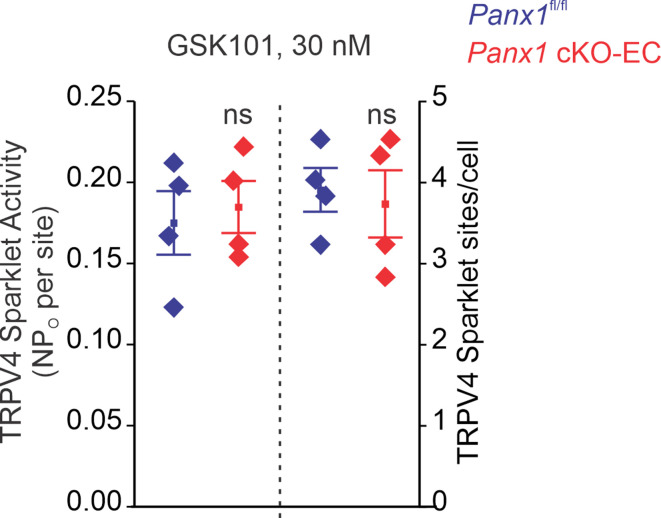
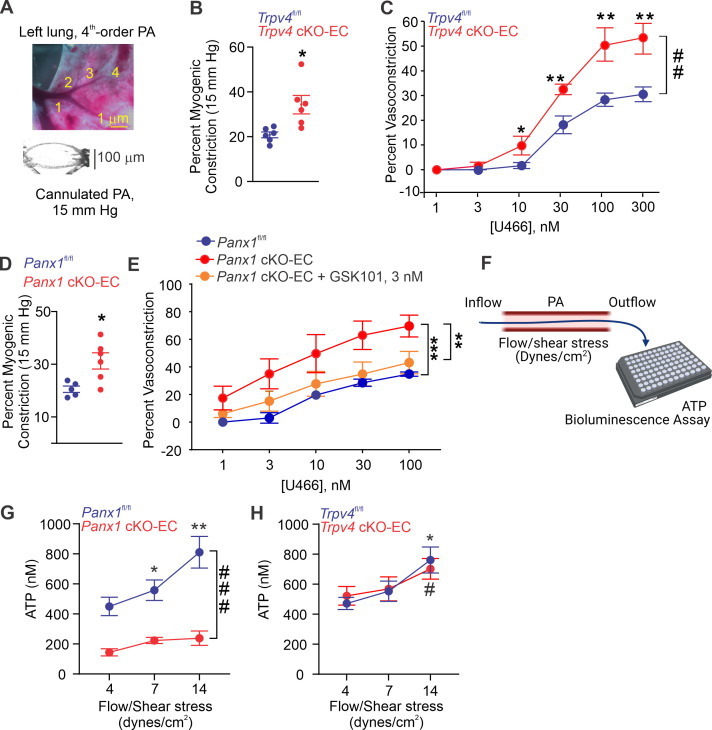
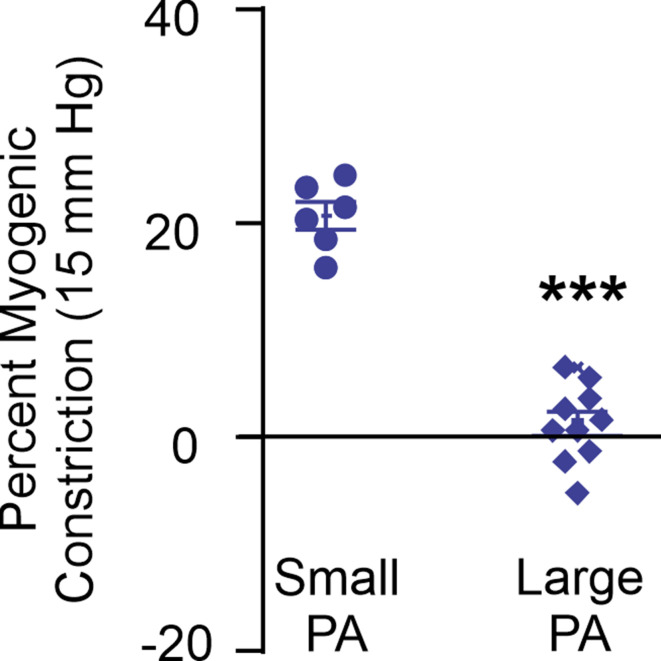
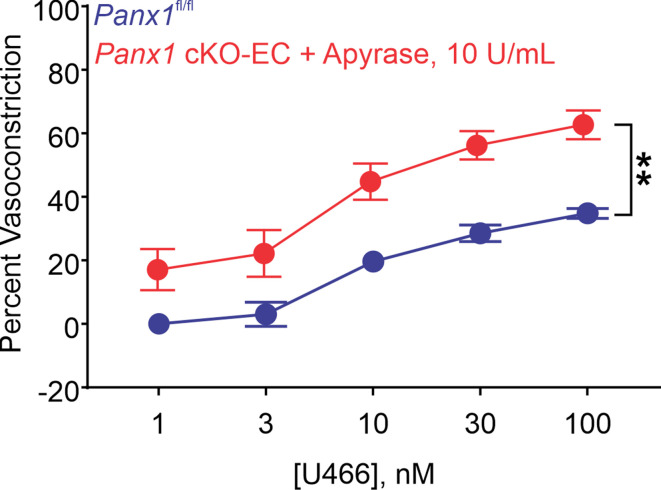
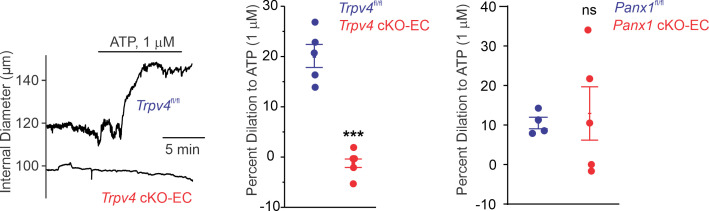
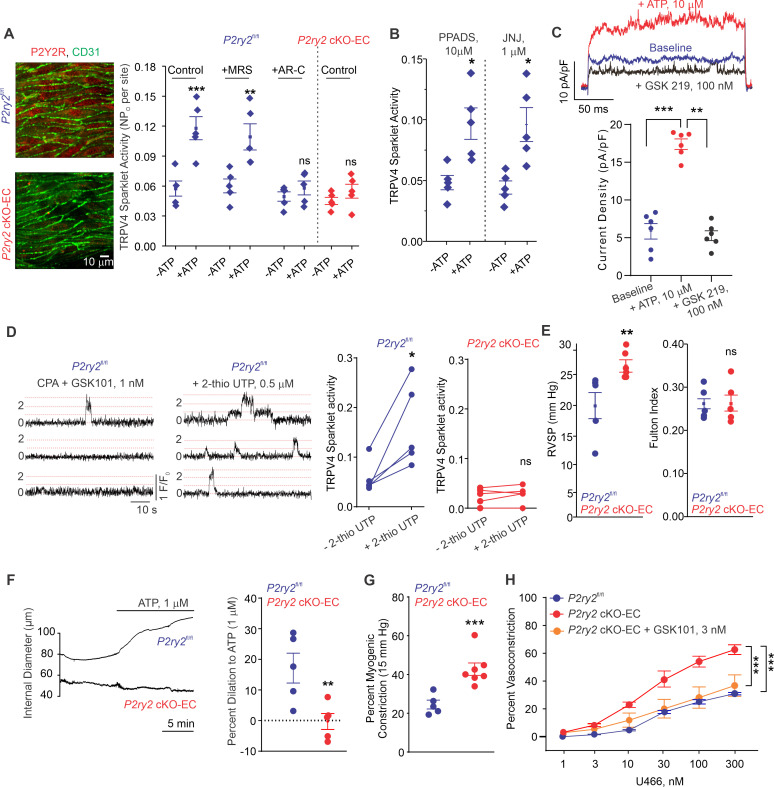
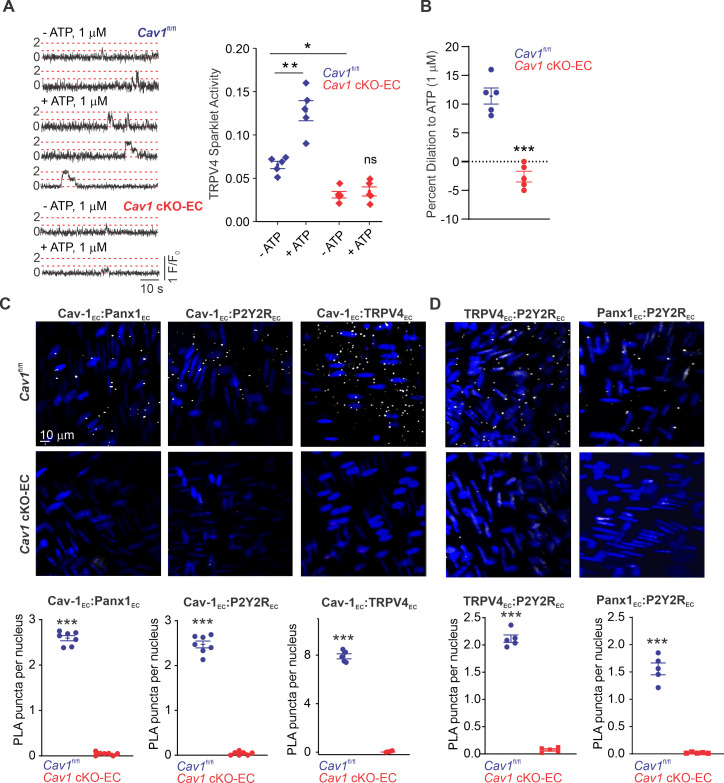
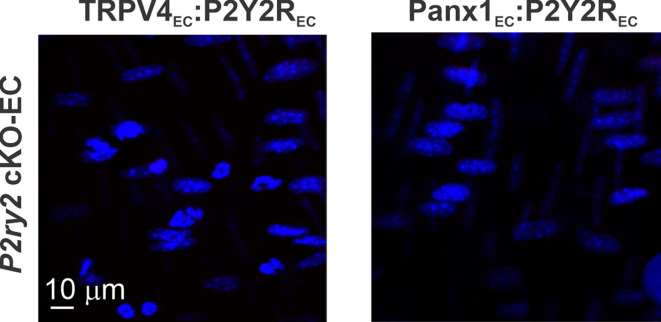
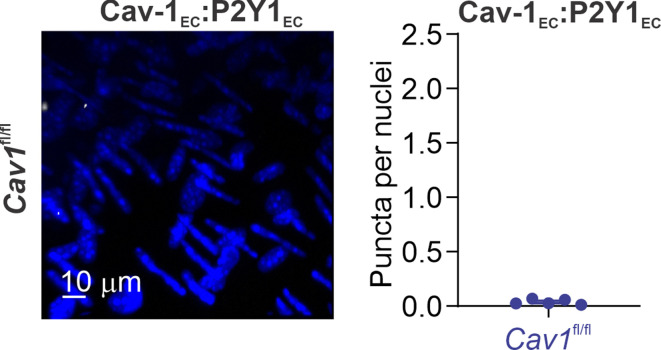
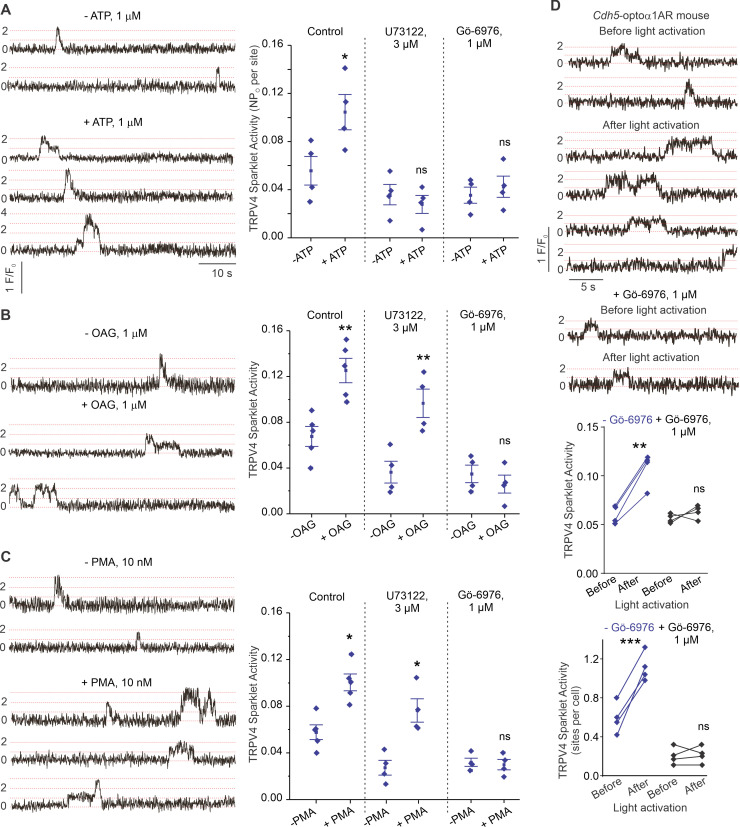
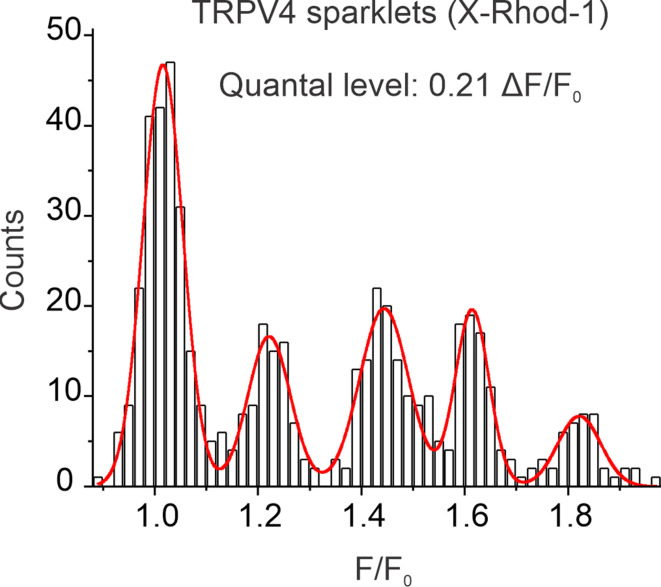
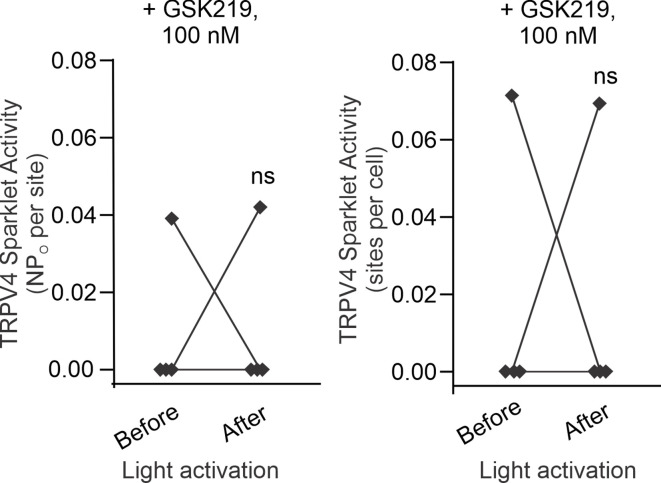
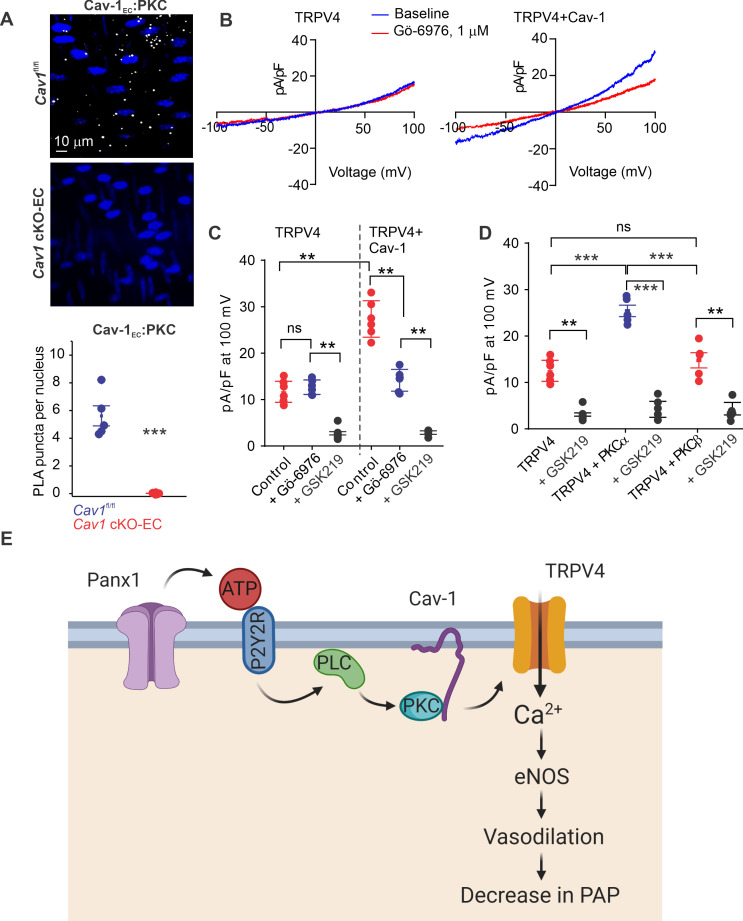
Pannexin 1 (Panx1), an ATP-efflux pathway, has been linked with inflammation in pulmonary capillaries. However, the physiological roles of endothelial Panx1 in the pulmonary vasculature are unknown. Endothelial transient receptor potential vanilloid 4 (TRPV4) channels lower pulmonary artery (PA) contractility and exogenous ATP activates endothelial TRPV4 channels. We hypothesized that endothelial Panx1-ATP-TRPV4 channel signaling promotes vasodilation and lowers pulmonary arterial pressure (PAP). Endothelial, but not smooth muscle, knockout of Panx1 increased PA contractility and raised PAP in mice. Flow/shear stress increased ATP efflux through endothelial Panx1 in PAs. Panx1-effluxed extracellular ATP signaled through purinergic P2Y2 receptor (P2Y2R) to activate protein kinase Cα (PKCα), which in turn activated endothelial TRPV4 channels. Finally, caveolin-1 provided a signaling scaffold for endothelial Panx1, P2Y2R, PKCα, and TRPV4 channels in PAs, promoting their spatial proximity and enabling signaling interactions. These results indicate that endothelial Panx1-P2Y2R-TRPV4 channel signaling, facilitated by caveolin-1, reduces PA contractility and lowers PAP in mice.
Keywords: Caveolin 1; Pannexin 1; TRP channel; biochemistry; cell biology; chemical biology; mouse; pulmonary vasculature; purinergic signaling.
© 2021, Daneva et al.
Conflict of interest statement
ZD, YC, EK, MK, SS, RM, CS, VL, BI, SS none, MO None
Figures
References
-
- Bakhshi FR, Mao M, Shajahan AN, Piegeler T, Chen Z, Chernaya O, Sharma T, Elliott WM, Szulcek R, Bogaard HJ, Comhair S, Erzurum S, van Nieuw Amerongen GP, Bonini MG, Minshall RD. Nitrosation-dependent caveolin 1 phosphorylation, ubiquitination, and degradation and its association with idiopathic pulmonary arterial hypertension. Pulmonary Circulation. 2013;3:816–830. doi: 10.1086/674753. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
