

Nitric Oxide Inhibition of Rickettsia rickettsii
- PMID: 34491789
- PMCID: PMC8594597
- DOI: 10.1128/IAI.00371-21
Nitric Oxide Inhibition of Rickettsia rickettsii
Abstract
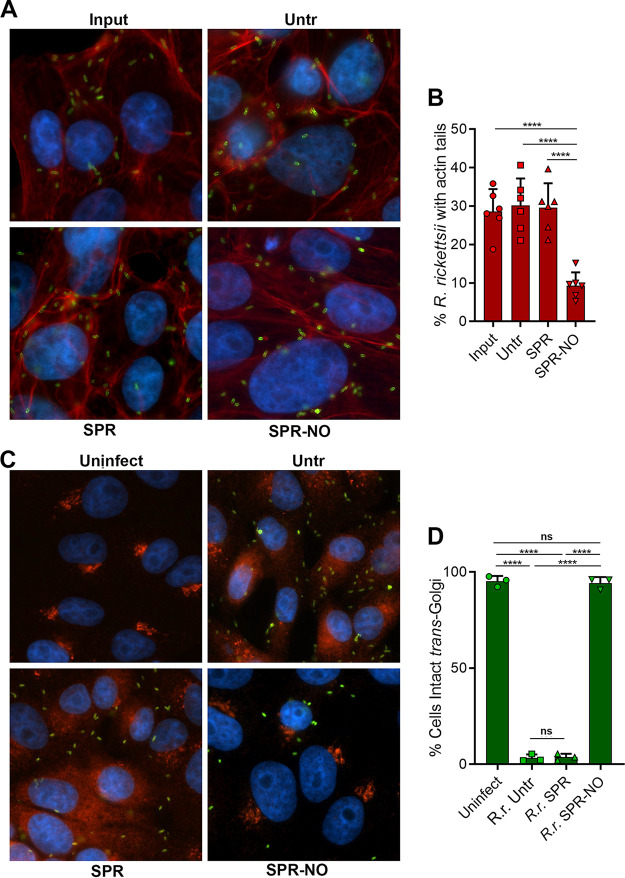
Rickettsia rickettsii, the causative agent of Rocky Mountain spotted fever, is an enzootic, obligate, intracellular bacterial pathogen. Nitric oxide (NO) synthesized by the inducible NO synthase (iNOS) is a potent antimicrobial component of innate immunity and has been implicated in the control of virulent Rickettsia spp. in diverse cell types. In this study, we examined the antibacterial role of NO on R. rickettsii. Our results indicate that NO challenge dramatically reduces R. rickettsii adhesion through the disruption of bacterial energetics. Additionally, NO-treated R. rickettsii cells were unable to synthesize protein or replicate in permissive cells. Activated, NO-producing macrophages restricted R. rickettsii infections, but inhibition of iNOS ablated the inhibition of bacterial growth. These data indicate that NO is a potent antirickettsial effector of innate immunity that targets energy generation in these pathogenic bacteria to prevent growth and subversion of infected host cells.
Keywords: Rickettsia; host defense; innate immunity; nitric oxide.
Figures
References
-
- Paddock CD, Denison AM, Lash RR, Liu L, Bollweg BC, Dahlgren FS, Kanamura CT, Angerami RN, Pereira dos Santos FC, Brasil Martines R, Karpathy SE. 2014. Phylogeograpy of Rickettsia rickettsii genotypes associated with fatal Rocky Mountain spotted fever. Am J Trop Med Hyg 91:589–597. 10.4269/ajtmh.14-0146. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
