

CREBH normalizes dyslipidemia and halts atherosclerosis in diabetes by decreasing circulating remnant lipoproteins
- PMID: 34491909
- PMCID: PMC8592537
- DOI: 10.1172/JCI153285
CREBH normalizes dyslipidemia and halts atherosclerosis in diabetes by decreasing circulating remnant lipoproteins
Abstract
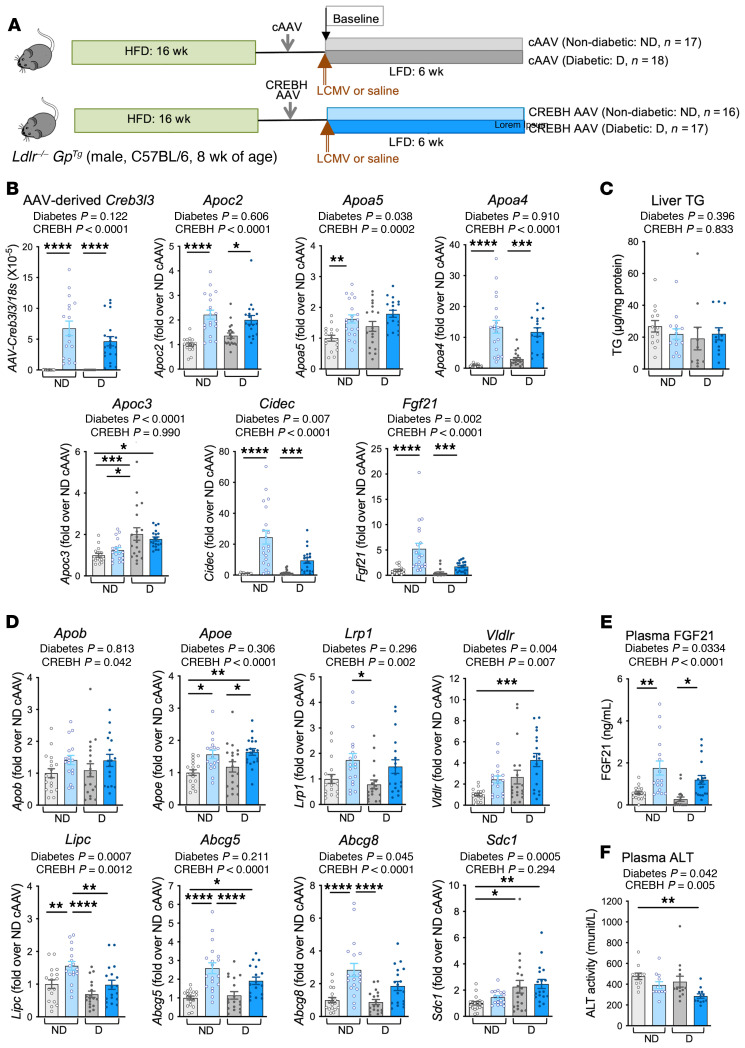
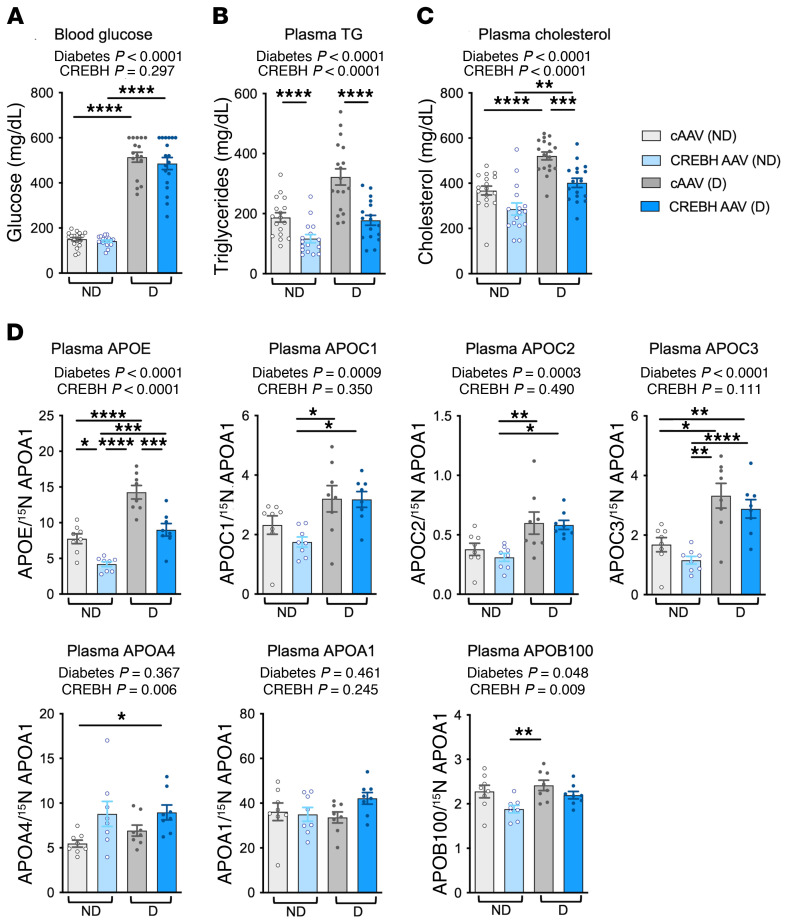
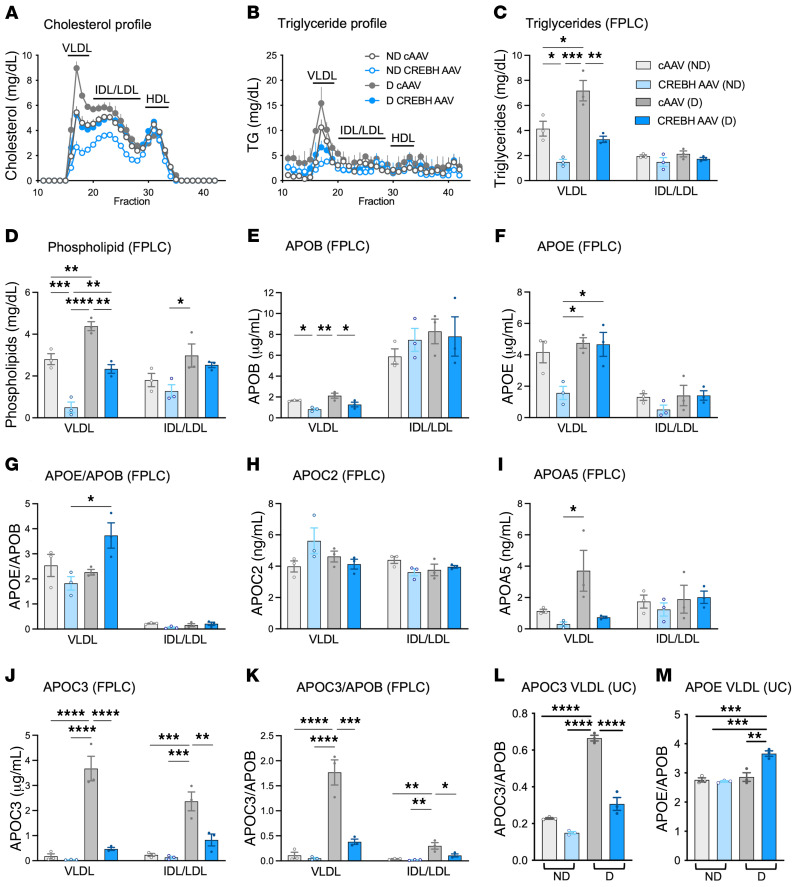
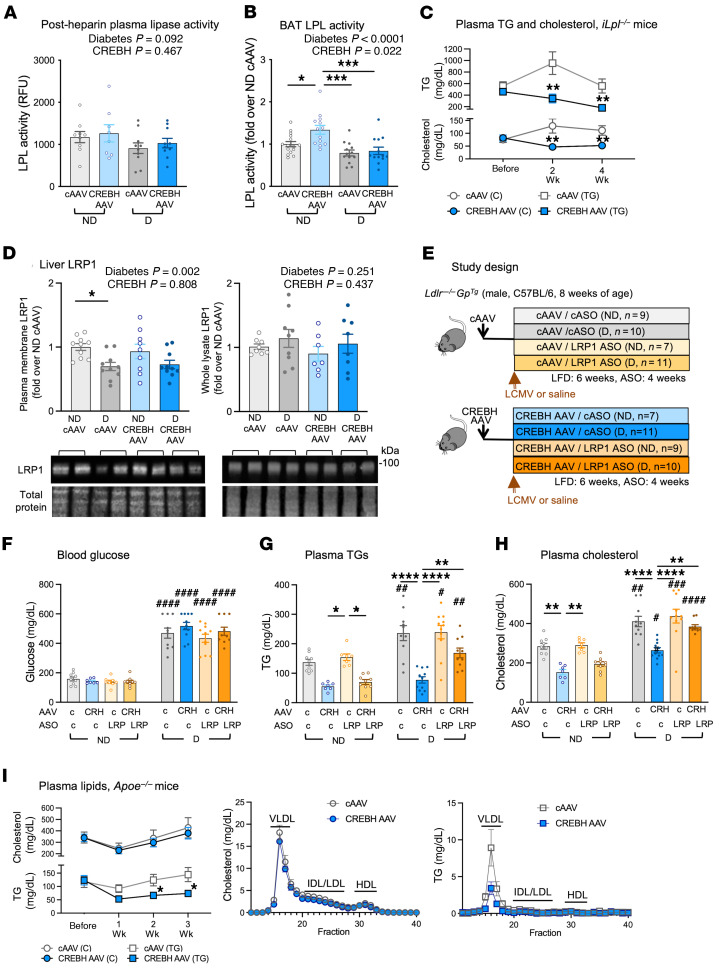
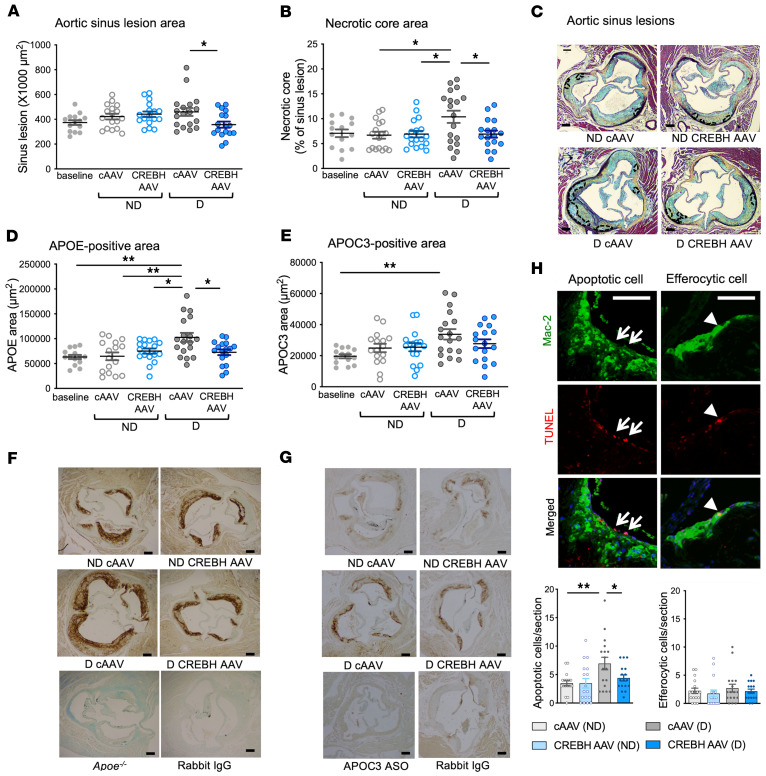
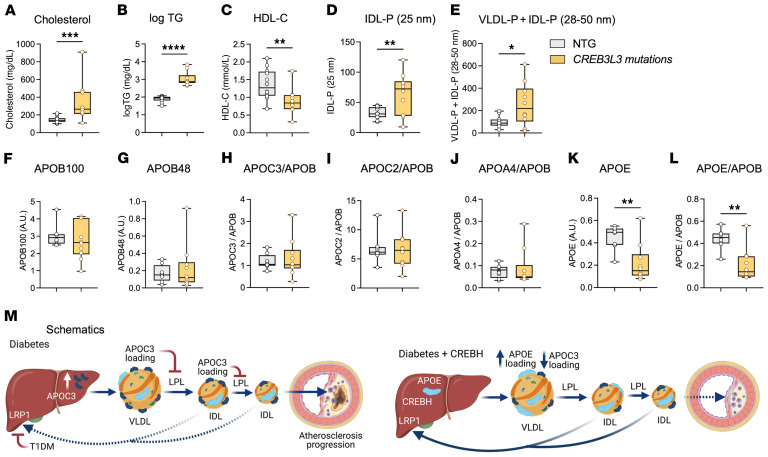
Loss-of-function mutations in the transcription factor CREB3L3 (CREBH) associate with severe hypertriglyceridemia in humans. CREBH is believed to lower plasma triglycerides by augmenting the activity of lipoprotein lipase (LPL). However, by using a mouse model of type 1 diabetes mellitus (T1DM), we found that greater liver expression of active CREBH normalized both elevated plasma triglycerides and cholesterol. Residual triglyceride-rich lipoprotein (TRL) remnants were enriched in apolipoprotein E (APOE) and impoverished in APOC3, an apolipoprotein composition indicative of increased hepatic clearance. The underlying mechanism was independent of LPL, as CREBH reduced both triglycerides and cholesterol in LPL-deficient mice. Instead, APOE was critical for CREBH's ability to lower circulating remnant lipoproteins because it failed to reduce TRL cholesterol in Apoe-/- mice. Importantly, individuals with CREB3L3 loss-of-function mutations exhibited increased levels of remnant lipoproteins that were deprived of APOE. Recent evidence suggests that impaired clearance of TRL remnants promotes cardiovascular disease in patients with T1DM. Consistently, we found that hepatic expression of CREBH prevented the progression of diabetes-accelerated atherosclerosis. Our results support the proposal that CREBH acts through an APOE-dependent pathway to increase hepatic clearance of remnant lipoproteins. They also implicate elevated levels of remnants in the pathogenesis of atherosclerosis in T1DM.
Keywords: Atherosclerosis; Diabetes; Endocrinology; Lipoproteins; Metabolism.
Conflict of interest statement
Figures
Comment in
-
Recruiting a transcription factor in the liver to prevent atherosclerosis.J Clin Invest. 2021 Nov 15;131(22):e154677. doi: 10.1172/JCI154677. J Clin Invest. 2021. PMID: 34779409 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
