

Lactate cross-talk in host-pathogen interactions
- PMID: 34492096
- PMCID: PMC8454702
- DOI: 10.1042/BCJ20210263
Lactate cross-talk in host-pathogen interactions
Abstract
Lactate is the main product generated at the end of anaerobic glycolysis or during the Warburg effect and its role as an active signalling molecule is increasingly recognised. Lactate can be released and used by host cells, by pathogens and commensal organisms, thus being essential for the homeostasis of host-microbe interactions. Infection can alter this intricate balance, and the presence of lactate transporters in most human cells including immune cells, as well as in a variety of pathogens (including bacteria, fungi and complex parasites) demonstrates the importance of this metabolite in regulating host-pathogen interactions. This review will cover lactate secretion and sensing in humans and microbes, and will discuss the existing evidence supporting a role for lactate in pathogen growth and persistence, together with lactate's ability to impact the orchestration of effective immune responses. The ubiquitous presence of lactate in the context of infection and the ability of both host cells and pathogens to sense and respond to it, makes manipulation of lactate a potential novel therapeutic strategy. Here, we will discuss the preliminary research that has been carried out in the context of cancer, autoimmunity and inflammation.
Keywords: host–pathogen interactions; immunometabolism; infection; lactate.
© 2021 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures
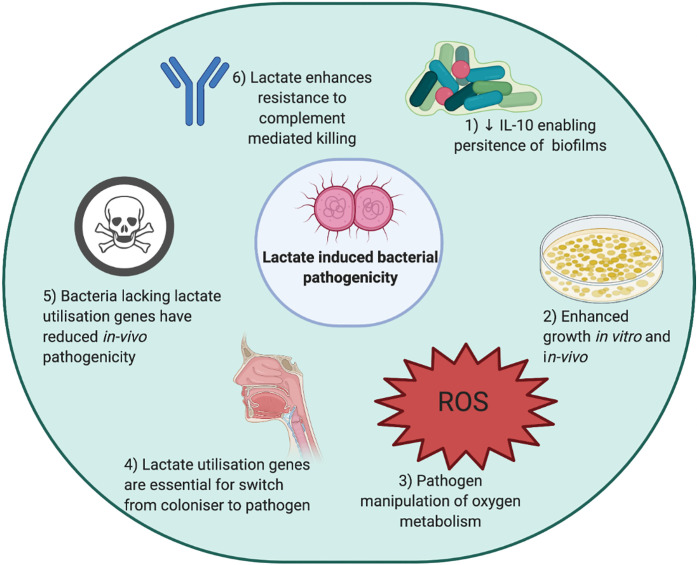
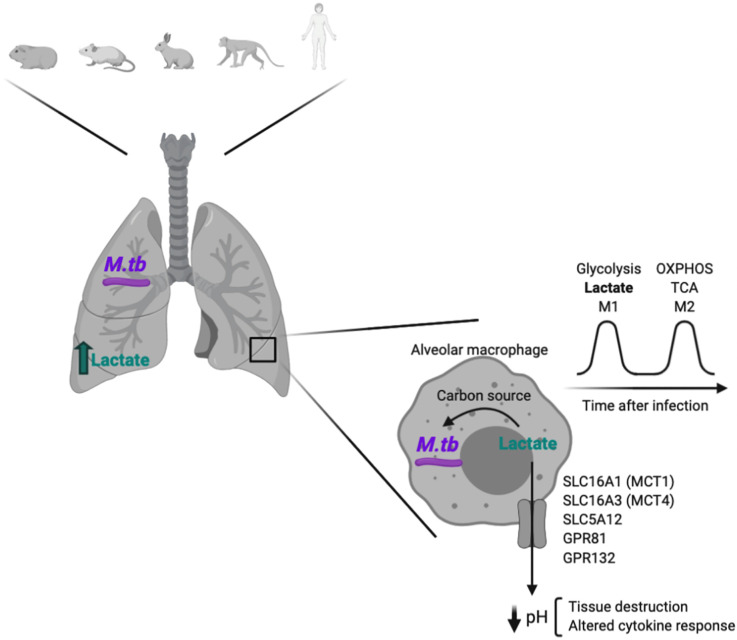
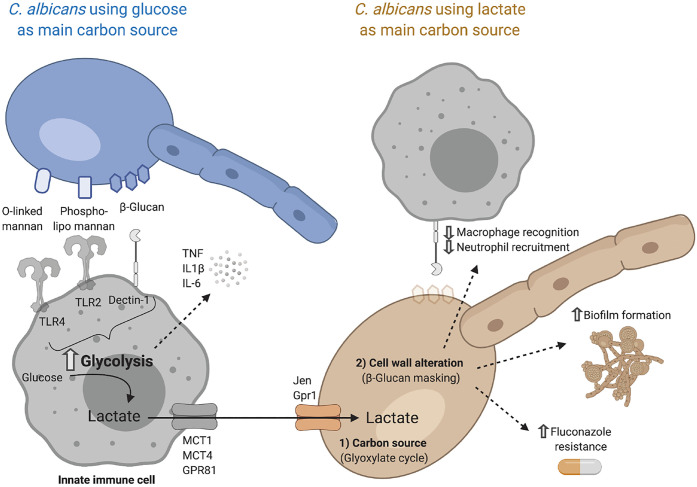
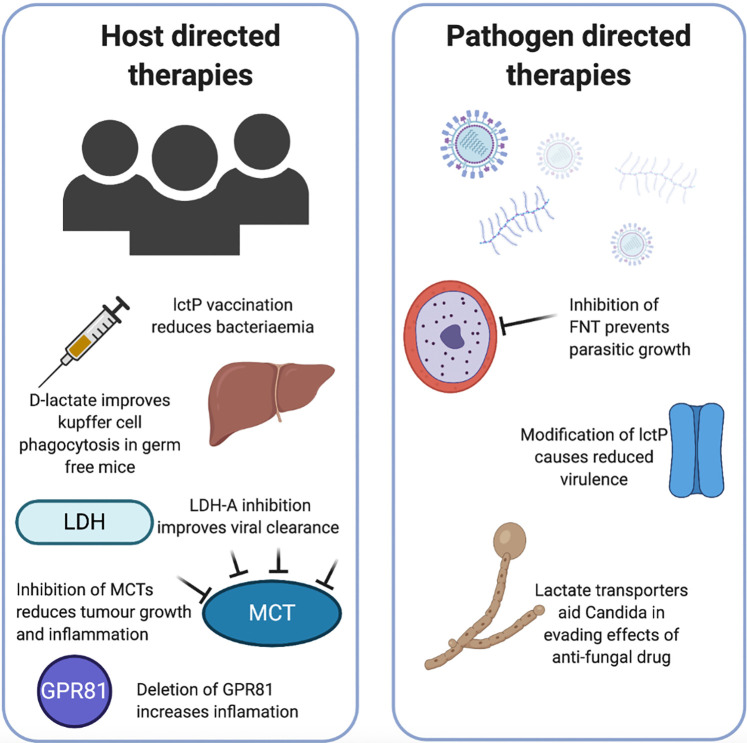
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases