

Gut microbiota regulation of P-glycoprotein in the intestinal epithelium in maintenance of homeostasis
- PMID: 34493329
- PMCID: PMC8425172
- DOI: 10.1186/s40168-021-01137-3
Gut microbiota regulation of P-glycoprotein in the intestinal epithelium in maintenance of homeostasis
Abstract
Background: P-glycoprotein (P-gp) plays a critical role in protection of the intestinal epithelia by mediating efflux of drugs/xenobiotics from the intestinal mucosa into the gut lumen. Recent studies bring to light that P-gp also confers a critical link in communication between intestinal mucosal barrier function and the innate immune system. Yet, despite knowledge for over 10 years that P-gp plays a central role in gastrointestinal homeostasis, the precise molecular mechanism that controls its functional expression and regulation remains unclear. Here, we assessed how the intestinal microbiome drives P-gp expression and function.
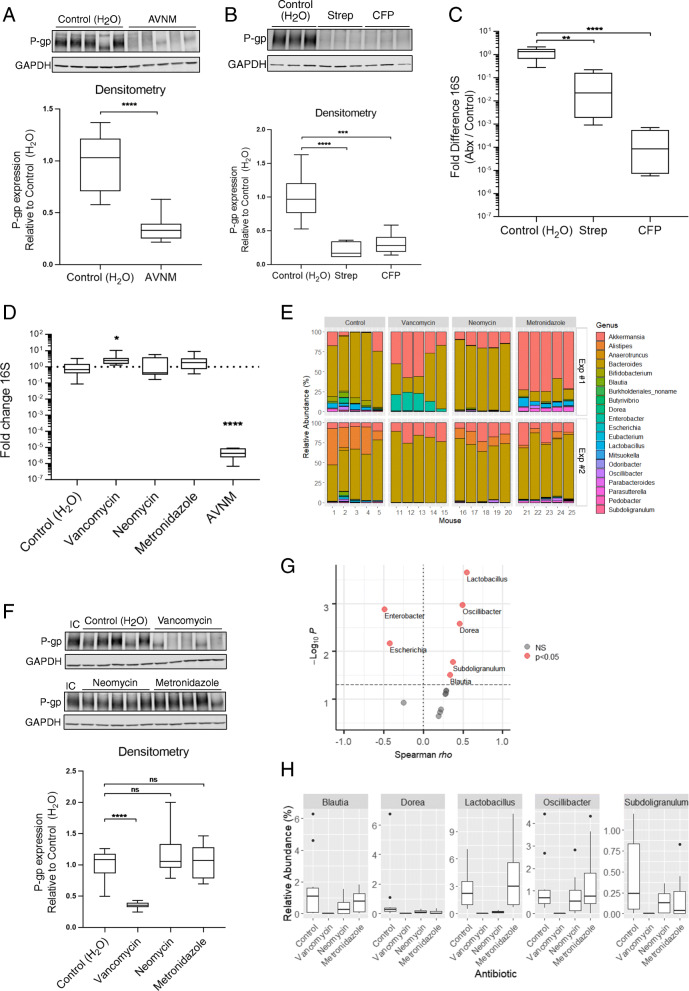
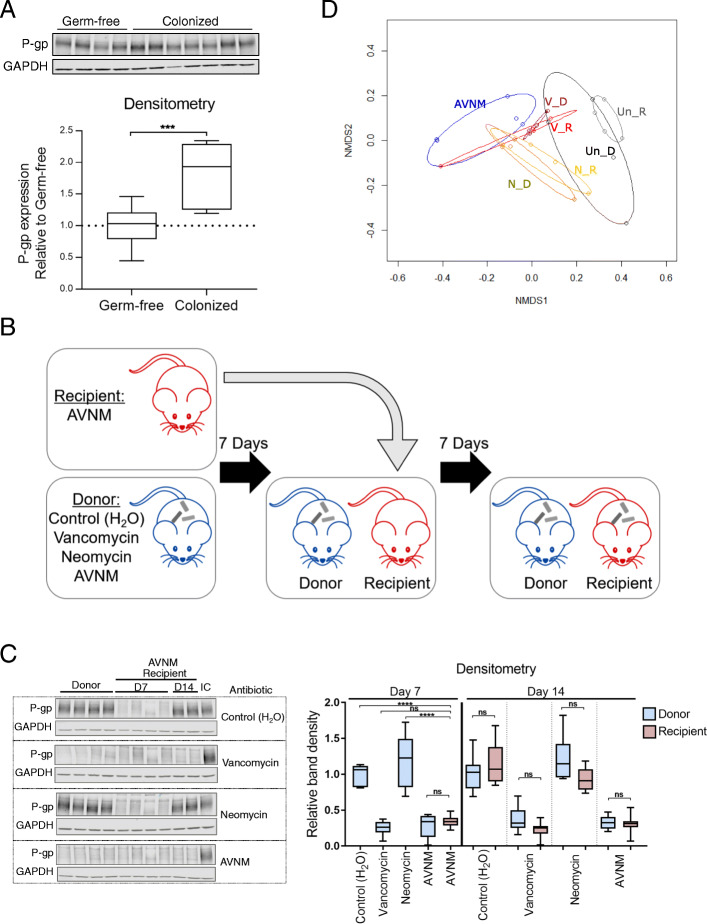
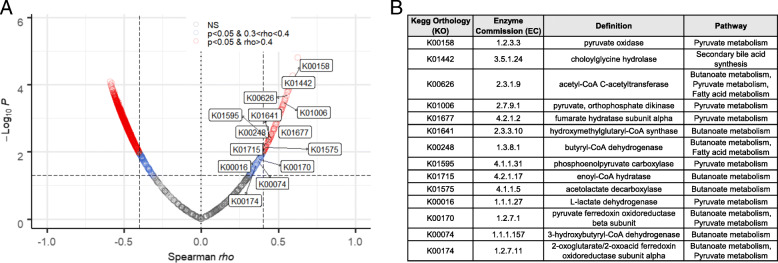
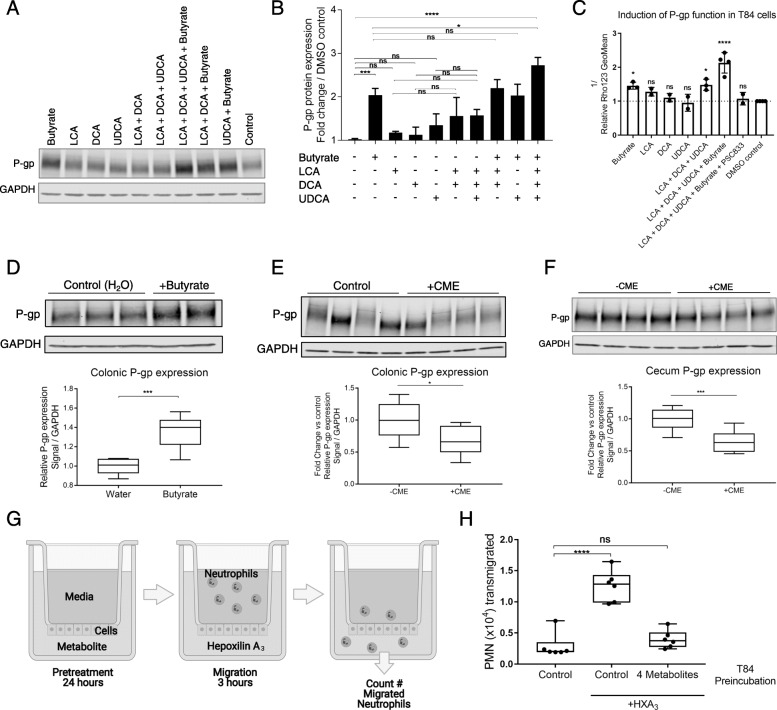
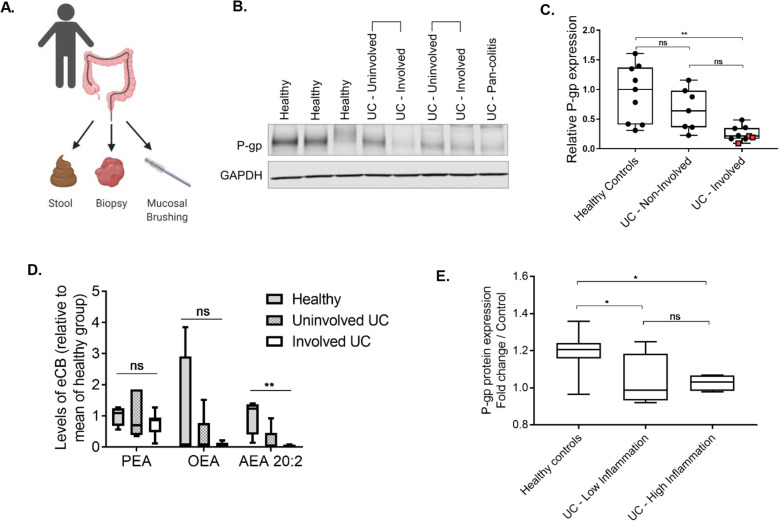
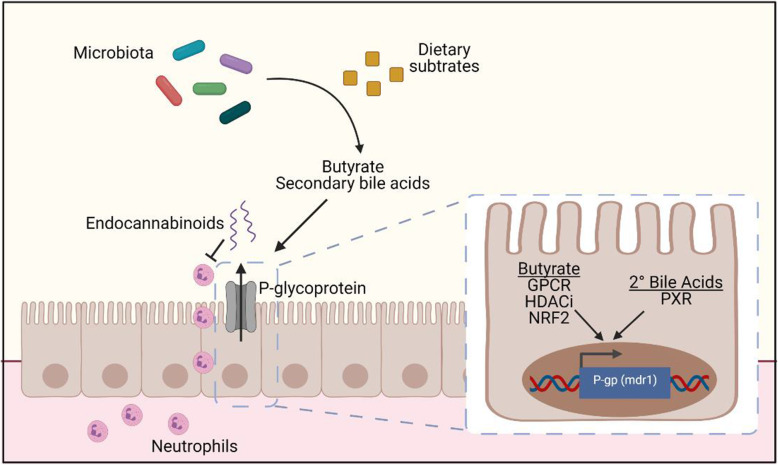
Results: We have identified a "functional core" microbiome of the intestinal gut community, specifically genera within the Clostridia and Bacilli classes, that is necessary and sufficient for P-gp induction in the intestinal epithelium in mouse models. Metagenomic analysis of this core microbial community revealed that short-chain fatty acid and secondary bile acid production positively associate with P-gp expression. We have further shown these two classes of microbiota-derived metabolites synergistically upregulate P-gp expression and function in vitro and in vivo. Moreover, in patients suffering from ulcerative colitis (UC), we find diminished P-gp expression coupled to the reduction of epithelial-derived anti-inflammatory endocannabinoids and luminal content (e.g., microbes or their metabolites) with a reduced capability to induce P-gp expression.
Conclusion: Overall, by means of both in vitro and in vivo studies as well as human subject sample analysis, we identify a mechanistic link between cooperative functional outputs of the complex microbial community and modulation of P-gp, an epithelial component, that functions to suppress overactive inflammation to maintain intestinal homeostasis. Hence, our data support a new cross-talk paradigm in microbiome regulation of mucosal inflammation. Video abstract.
Keywords: Endocannabinoid; Inflammation; Inflammatory bowel diseases; Intestinal epithelium; Microbiome; Multi-drug resistance transporter; P-glycoprotein; Secondary bile acids; Short-chain fatty acids; Ulcerative colitis.
© 2021. The Author(s).
Conflict of interest statement
B.A.M. and R.J.M. are coinventors on a patent application (PGT/US 18/42116) emanating, in part, from the findings described herein. They, along with their respective academic institutions, stand to gain financially through potential commercialization outcomes resulting from activities associated with the licensing of that intellectual property.
Figures
References
-
- Pazos M, Siccardi D, Mumy KL, Bien JD, Louie S, Shi HN, Gronert K, Mrsny RJ, McCormick BA. Multidrug resistance-associated transporter 2 regulates mucosal inflammation by facilitating the synthesis of hepoxilin A3. J Immunol. 2008;181(11):8044–8052. doi: 10.4049/jimmunol.181.11.8044. - DOI - PMC - PubMed
-
- Szabady RL, Louissaint C, Lubben A, Xie B, Reeksting S, Tuohy C, Demma Z, Foley SE, Faherty CS, Llanos-Chea A, Olive AJ, Mrsny RJ, McCormick BA. Intestinal P-glycoprotein exports endocannabinoids to prevent inflammation and maintain homeostasis. J Clin Invest. 2018;128(9):4044–4056. doi: 10.1172/JCI96817. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
