

5'-Cap sequestration is an essential determinant of HIV-1 genome packaging
- PMID: 34493679
- PMCID: PMC8449379
- DOI: 10.1073/pnas.2112475118
5'-Cap sequestration is an essential determinant of HIV-1 genome packaging
Abstract
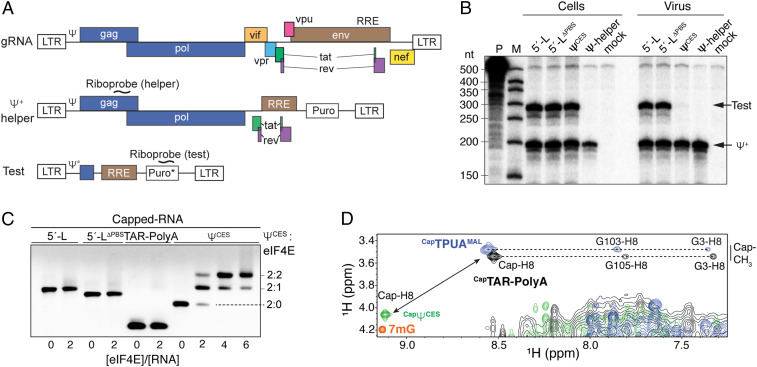
HIV-1 selectively packages two copies of its 5'-capped RNA genome (gRNA) during virus assembly, a process mediated by the nucleocapsid (NC) domain of the viral Gag polyprotein and encapsidation signals located within the dimeric 5' leader of the viral RNA. Although residues within the leader that promote packaging have been identified, the determinants of authentic packaging fidelity and efficiency remain unknown. Here, we show that a previously characterized 159-nt region of the leader that possesses all elements required for RNA dimerization, high-affinity NC binding, and packaging in a noncompetitive RNA packaging assay (ΨCES) is unexpectedly poorly packaged when assayed in competition with the intact 5' leader. ΨCES lacks a 5'-tandem hairpin element that sequesters the 5' cap, suggesting that cap sequestration may be important for packaging. Consistent with this hypothesis, mutations within the intact leader that expose the cap without disrupting RNA structure or NC binding abrogated RNA packaging, and genetic addition of a 5' ribozyme to ΨCES to enable cotranscriptional shedding of the 5' cap promoted ΨCES-mediated RNA packaging to wild-type levels. Additional mutations that either block dimerization or eliminate subsets of NC binding sites substantially attenuated competitive packaging. Our studies indicate that packaging is achieved by a bipartite mechanism that requires both sequestration of the 5' cap and exposure of NC binding sites that reside fully within the ΨCES region of the dimeric leader. We speculate that cap sequestration prevents irreversible capture by the cellular RNA processing and translation machinery, a mechanism likely employed by other viruses that package 5'-capped RNA genomes.
Keywords: 5′ cap; HIV-1; RNA; genome; packaging.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures




Comment in
-
Show your cap or be packaged into HIV-1.Proc Natl Acad Sci U S A. 2021 Oct 5;118(40):e2115344118. doi: 10.1073/pnas.2115344118. Proc Natl Acad Sci U S A. 2021. PMID: 34583998 Free PMC article. No abstract available.
References
-
- Coffin J. M., Hughes S. H., Varmus H. E., Retroviruses (Cold Spring Harbor Laboratory Press, Plainview, NY, 1997). - PubMed
-
- Berkowitz R., Fisher J., Goff S. P., RNA packaging. Curr. Top. Microbiol. Immunol. 214, 177–218 (1996). - PubMed
-
- Paillart J.-C., Shehu-Xhilaga M., Marquet R., Mak J., Dimerization of retroviral RNA genomes: An inseparable pair. Nat. Rev. Microbiol. 2, 461–472 (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
