

Specific impact of β5t on proteasome subunit composition in cortical thymic epithelial cells
- PMID: 34496235
- PMCID: PMC8442848
- DOI: 10.1016/j.celrep.2021.109657
Specific impact of β5t on proteasome subunit composition in cortical thymic epithelial cells
Abstract
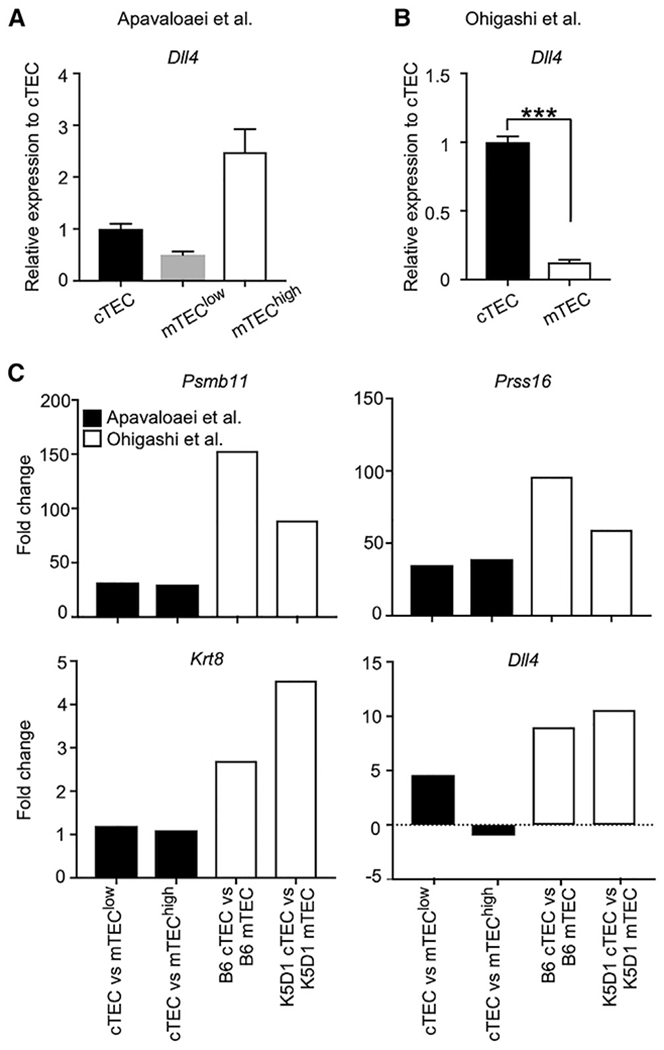
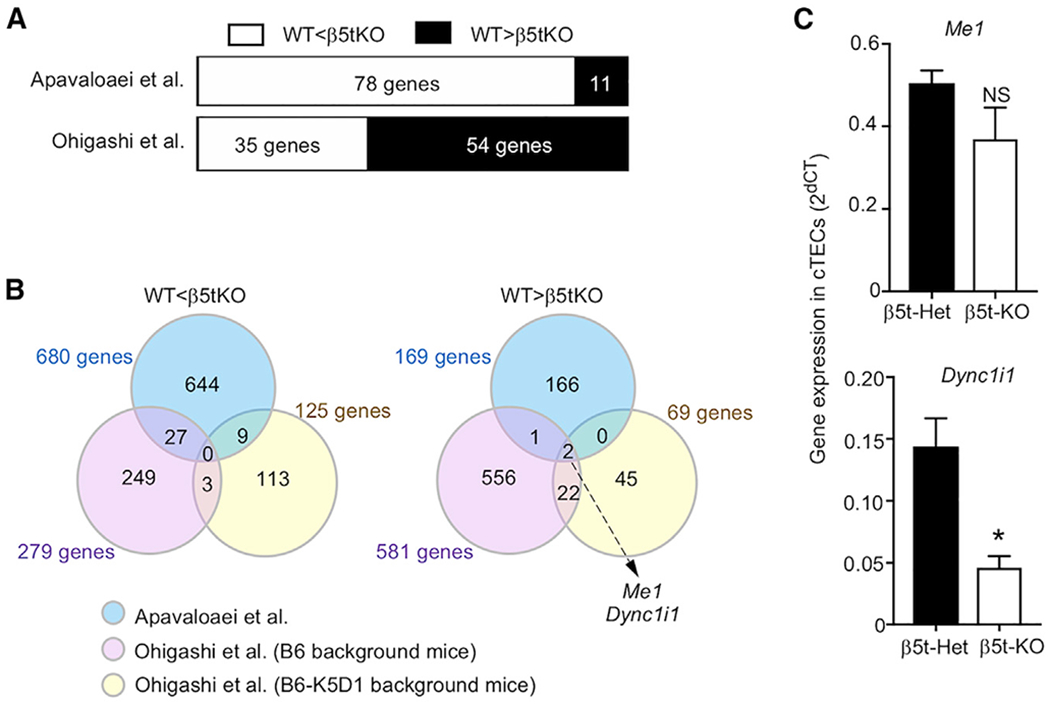
β5t is a cortical thymic epithelial cell (cTEC)-specific component of the thymoproteasome, which is essential for the optimal production of functionally competent CD8+ T cells. Our recent analysis showed a specific impact of β5t on proteasome subunit composition in cTECs, supporting the possibility that the thymoproteasome optimizes CD8+ T cell development through the production of MHC-I-associated unique self-peptides in cTECs. However, a recent article reports that β5t regulates the expression of hundreds of cTEC genes and affects both CD4+ and CD8+ thymocytes by causing oxidative stress in thymocytes. The authors further analyze our published data and describe that they confirm their conclusions. Here, we examine the issues that they raise and conclude that, rather than regulating hundreds of genes in cTECs, β5t has a highly specific impact in cTECs on proteasome subunit composition. This Matters Arising Response article addresses the Apavaloaei et al. (2021) Matters Arising paper, published concurrently in Cell Reports.
Keywords: Cortical thymic epithelial cell; Positive selection; Proteasome; Psmb11; T cell development; Thymoproteasome; Thymus; β5t.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

Comment on
-
Trans-omics Impact of Thymoproteasome in Cortical Thymic Epithelial Cells.Cell Rep. 2019 Nov 26;29(9):2901-2916.e6. doi: 10.1016/j.celrep.2019.10.079. Cell Rep. 2019. PMID: 31775054 Free PMC article.
-
PSMB11 regulates gene expression in cortical thymic epithelial cells.Cell Rep. 2021 Sep 7;36(10):109546. doi: 10.1016/j.celrep.2021.109546. Cell Rep. 2021. PMID: 34496243
References
-
- Apavaloaei A, Brochu S, Dong M, Rouette A, Hardy MP, Villafano G, Murata S, Melichar HJ, and Perreault C (2019). PSMB11 orchestrates the development of CD4 and CD8 thymocytes via regulation of gene expression in cortical thymic epithelial cells. J. Immunol 202, 966–978. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
