

Negative charge in the RACK1 loop broadens the translational capacity of the human ribosome
- PMID: 34496247
- PMCID: PMC8451006
- DOI: 10.1016/j.celrep.2021.109663
Negative charge in the RACK1 loop broadens the translational capacity of the human ribosome
Abstract
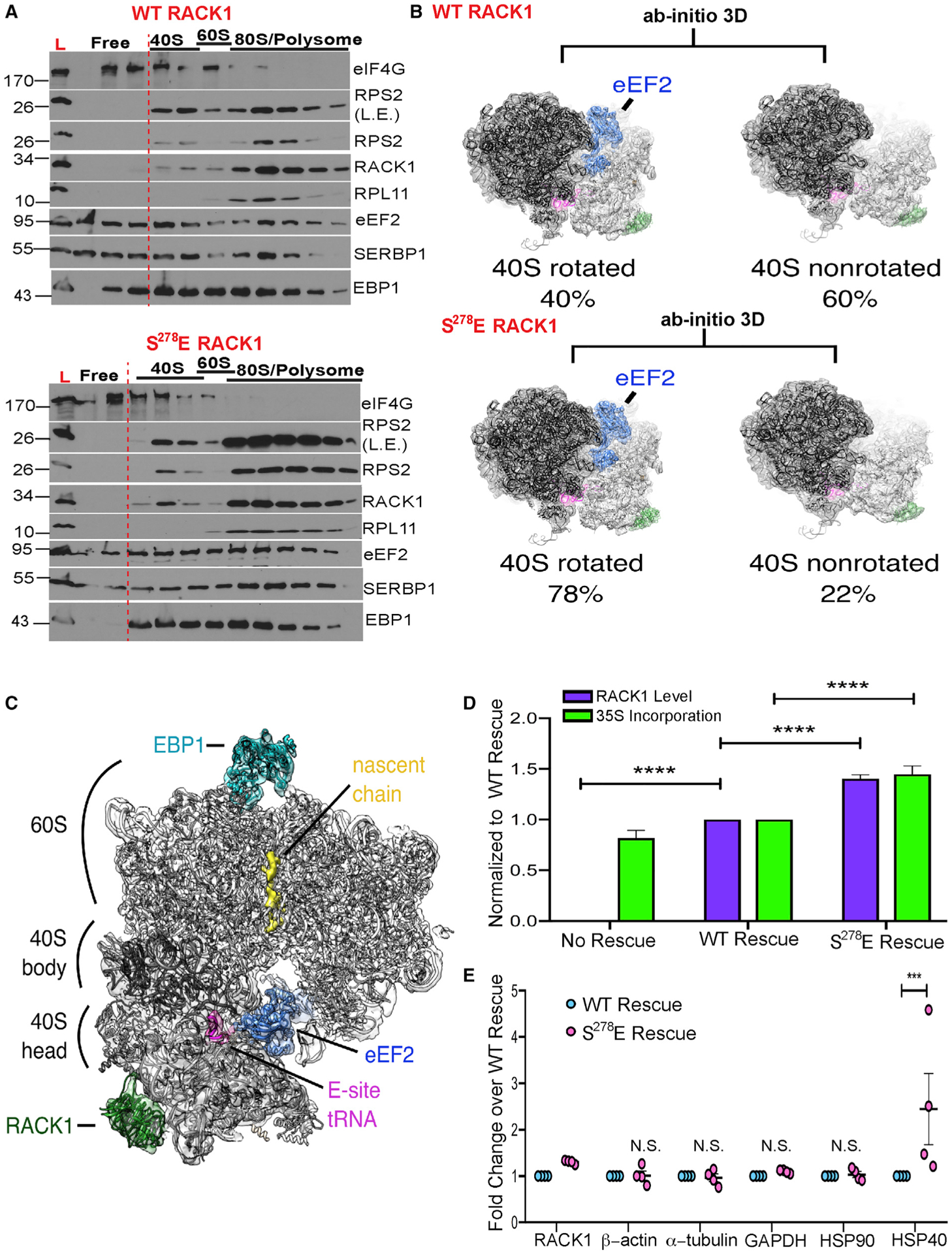
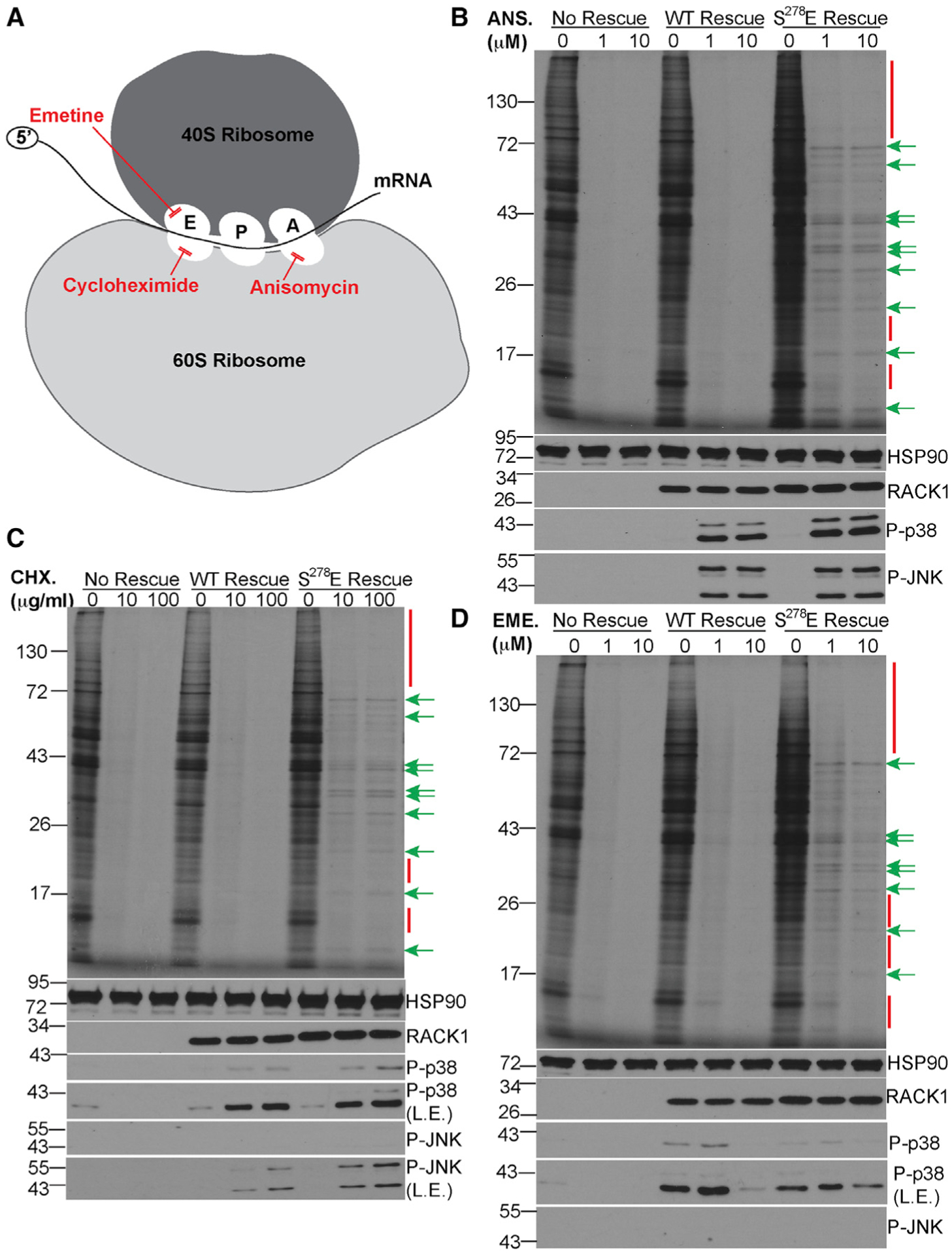
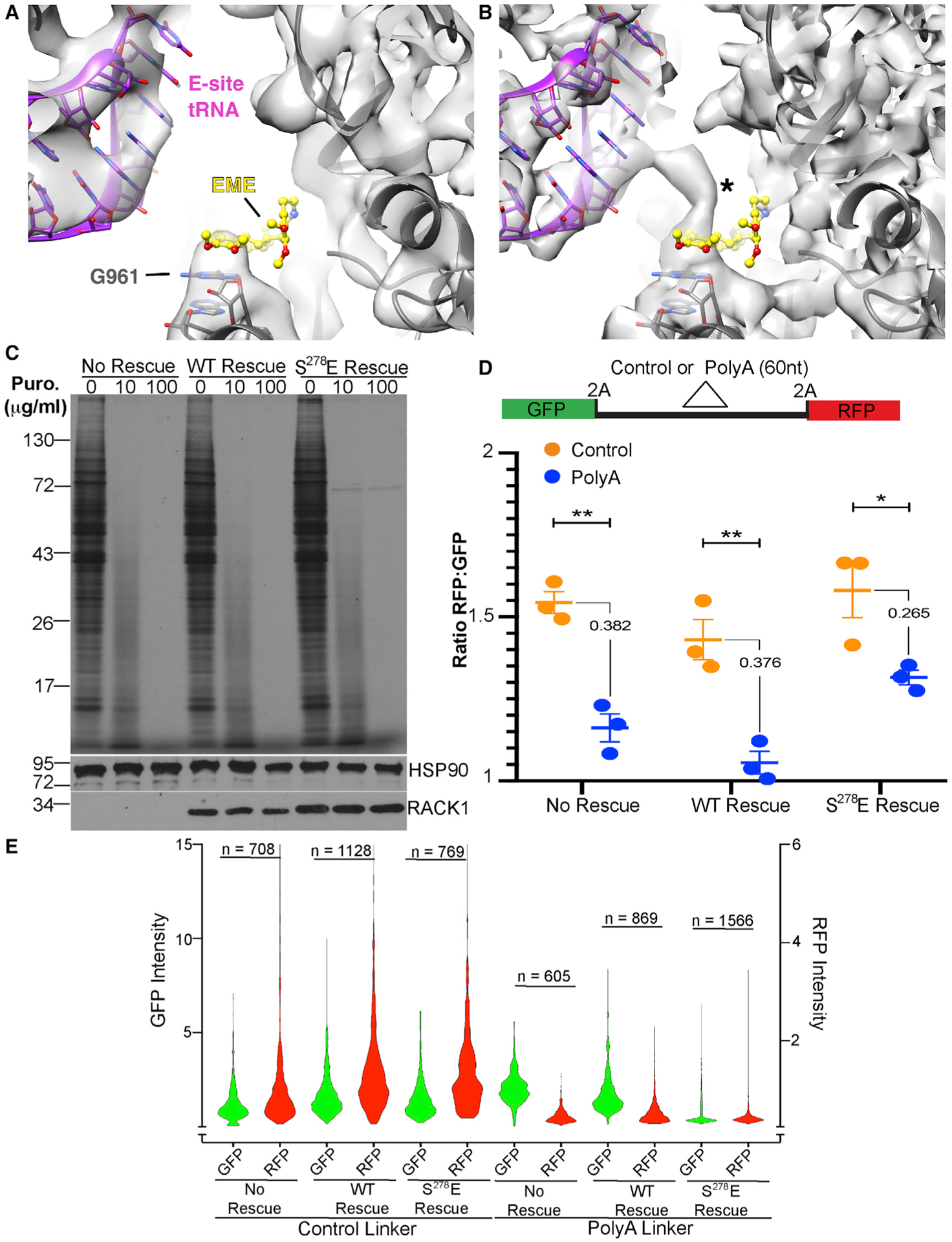
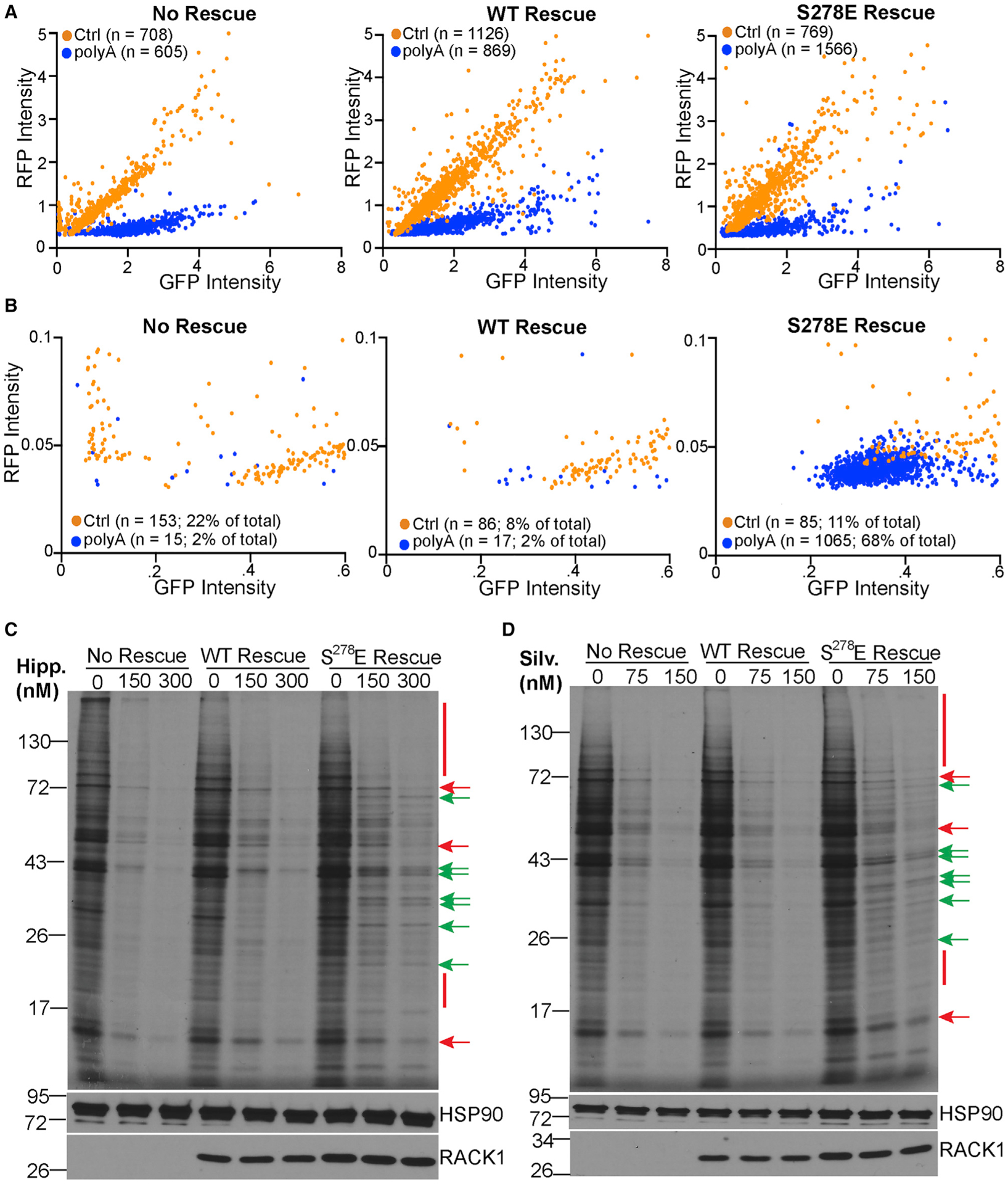
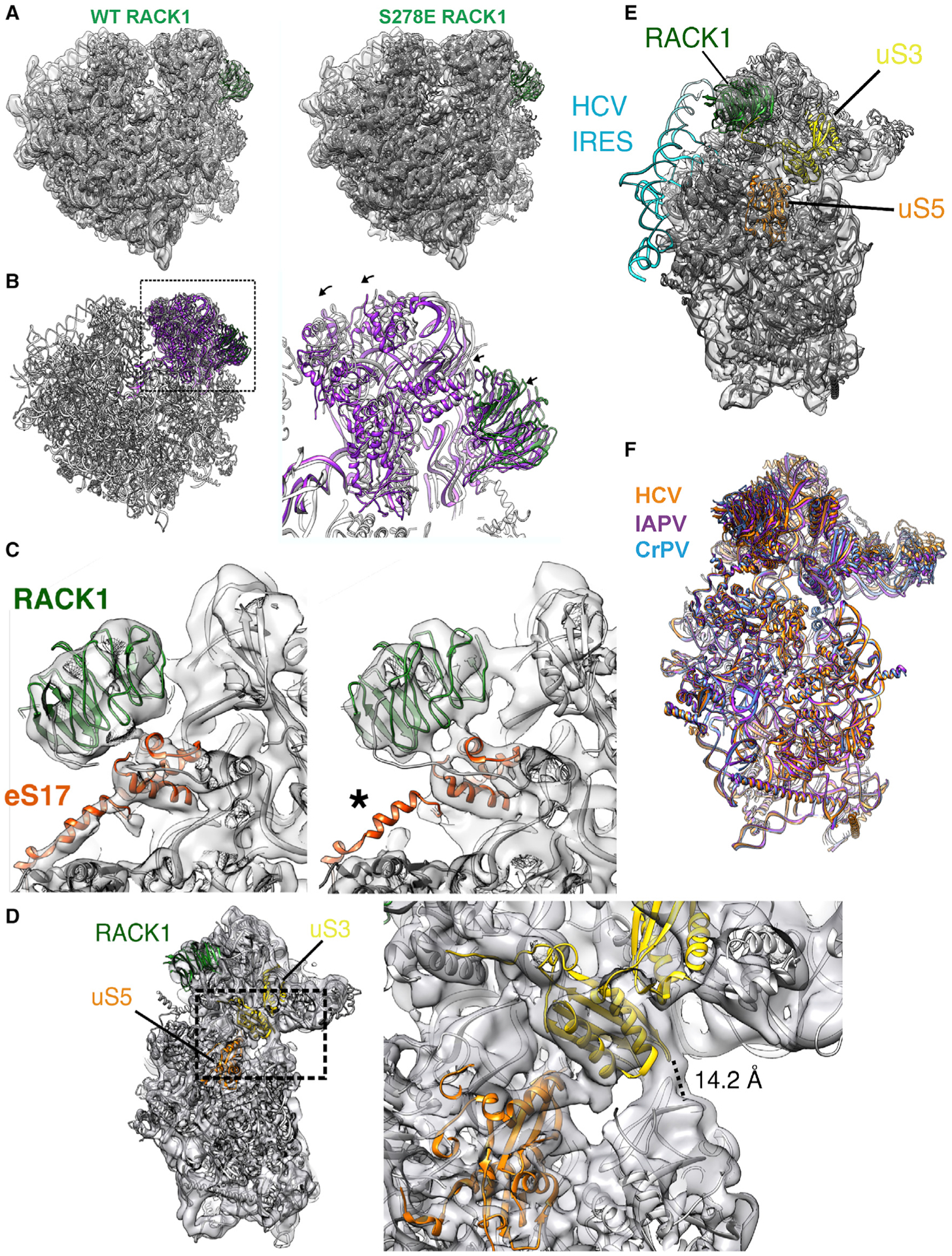
Although the roles of initiation factors, RNA binding proteins, and RNA elements in regulating translation are well defined, how the ribosome functionally diversifies remains poorly understood. In their human hosts, poxviruses phosphorylate serine 278 (S278) at the tip of a loop domain in the small subunit ribosomal protein RACK1, thereby mimicking negatively charged residues in the RACK1 loops of dicot plants and protists to stimulate translation of transcripts with 5' poly(A) leaders. However, how a negatively charged RACK1 loop affects ribosome structure and its broader translational output is not known. Here, we show that although ribotoxin-induced stress signaling and stalling on poly(A) sequences are unaffected, negative charge in the RACK1 loop alters the swivel motion of the 40S head domain in a manner similar to several internal ribosome entry sites (IRESs), confers resistance to various protein synthesis inhibitors, and broadly supports noncanonical modes of translation.
Keywords: IRES; alternative initiation; cryo-EM; mRNA specification; post-translational modification; protein synthesis; ribosome; selective translation; structure.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Trans-kingdom mimicry underlies ribosome customization by a poxvirus kinase.Nature. 2017 Jun 29;546(7660):651-655. doi: 10.1038/nature22814. Epub 2017 Jun 21. Nature. 2017. PMID: 28636603 Free PMC article.
-
RACK1 evolved species-specific multifunctionality in translational control through sequence plasticity within a loop domain.J Cell Sci. 2019 Jun 19;132(12):jcs228908. doi: 10.1242/jcs.228908. J Cell Sci. 2019. PMID: 31118235 Free PMC article.
-
Ribosomal RACK1:Protein Kinase C βII Phosphorylates Eukaryotic Initiation Factor 4G1 at S1093 To Modulate Cap-Dependent and -Independent Translation Initiation.Mol Cell Biol. 2018 Sep 14;38(19):e00304-18. doi: 10.1128/MCB.00304-18. Print 2018 Oct 1. Mol Cell Biol. 2018. PMID: 30012863 Free PMC article.
-
Hepatitis C Virus Translation Regulation.Int J Mol Sci. 2020 Mar 27;21(7):2328. doi: 10.3390/ijms21072328. Int J Mol Sci. 2020. PMID: 32230899 Free PMC article. Review.
-
Regulation of eukaryotic translation by the RACK1 protein: a platform for signalling molecules on the ribosome.EMBO Rep. 2004 Dec;5(12):1137-41. doi: 10.1038/sj.embor.7400291. EMBO Rep. 2004. PMID: 15577927 Free PMC article. Review.
Cited by
-
Proteostasis Deregulation in Neurodegeneration and Its Link with Stress Granules: Focus on the Scaffold and Ribosomal Protein RACK1.Cells. 2022 Aug 19;11(16):2590. doi: 10.3390/cells11162590. Cells. 2022. PMID: 36010666 Free PMC article. Review.
-
Ribosome customization and functional diversification among P-stalk proteins regulate late poxvirus protein synthesis.Cell Rep. 2025 Jan 28;44(1):115119. doi: 10.1016/j.celrep.2024.115119. Epub 2025 Jan 8. Cell Rep. 2025. PMID: 39786991 Free PMC article.
-
Ribosomes in poxvirus infection.Curr Opin Virol. 2022 Oct;56:101256. doi: 10.1016/j.coviro.2022.101256. Epub 2022 Oct 18. Curr Opin Virol. 2022. PMID: 36270183 Free PMC article. Review.
-
The role of ribosomal protein networks in ribosome dynamics.Nucleic Acids Res. 2025 Jan 7;53(1):gkae1308. doi: 10.1093/nar/gkae1308. Nucleic Acids Res. 2025. PMID: 39788545 Free PMC article.
-
Impacts of ribosomal RNA sequence variation on gene expression and phenotype.Philos Trans R Soc Lond B Biol Sci. 2025 Mar 6;380(1921):20230379. doi: 10.1098/rstb.2023.0379. Epub 2025 Mar 6. Philos Trans R Soc Lond B Biol Sci. 2025. PMID: 40045785 Free PMC article. Review.
References
-
- Anger AM, Armache JP, Berninghausen O, Habeck M, Subklewe M, Wilson DN, and Beckmann R (2013). Structures of the human and Drosophila 80S ribosome. Nature 497, 80–85. - PubMed
-
- Asarnow D, Palovcak E, and Cheng Y (2019). UCSF pyem v0.5. Zenodo.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
