

Chronic stress primes innate immune responses in mice and humans
- PMID: 34496250
- PMCID: PMC8493594
- DOI: 10.1016/j.celrep.2021.109595
Chronic stress primes innate immune responses in mice and humans
Abstract
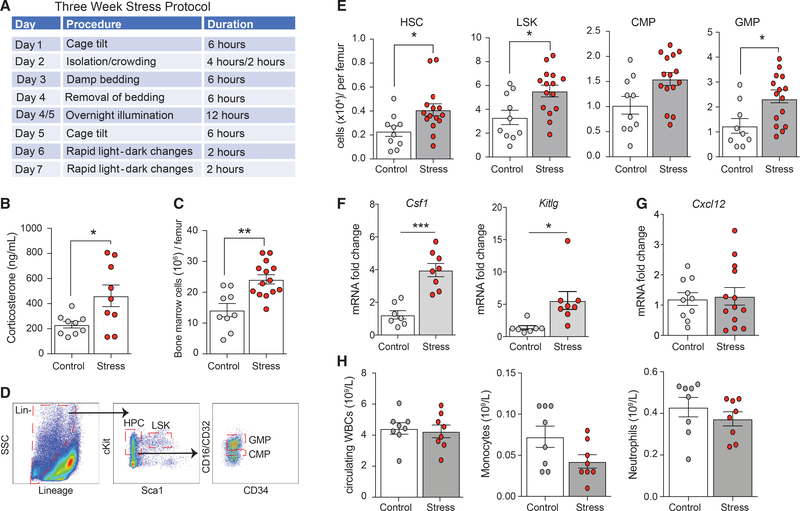
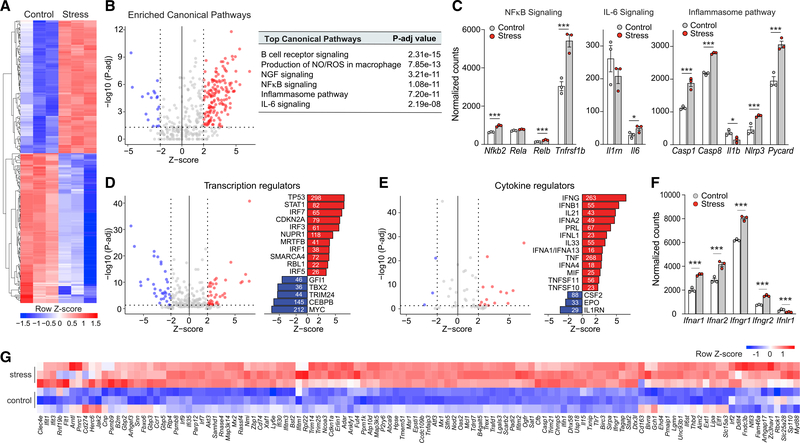
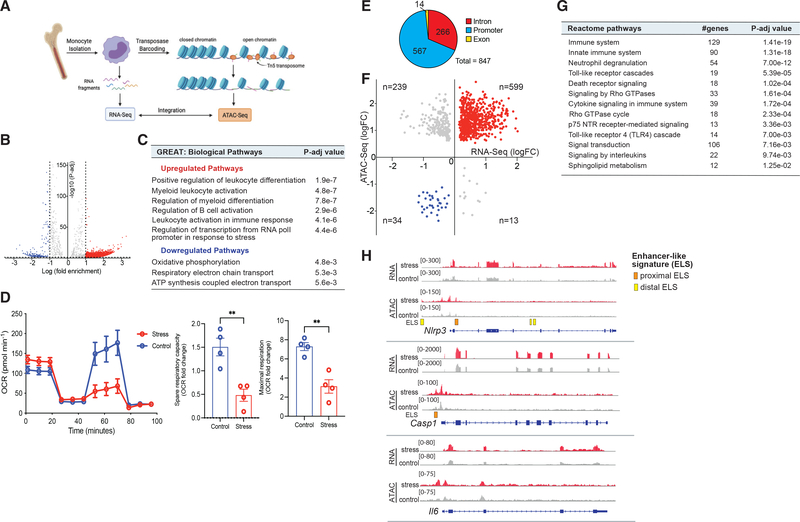
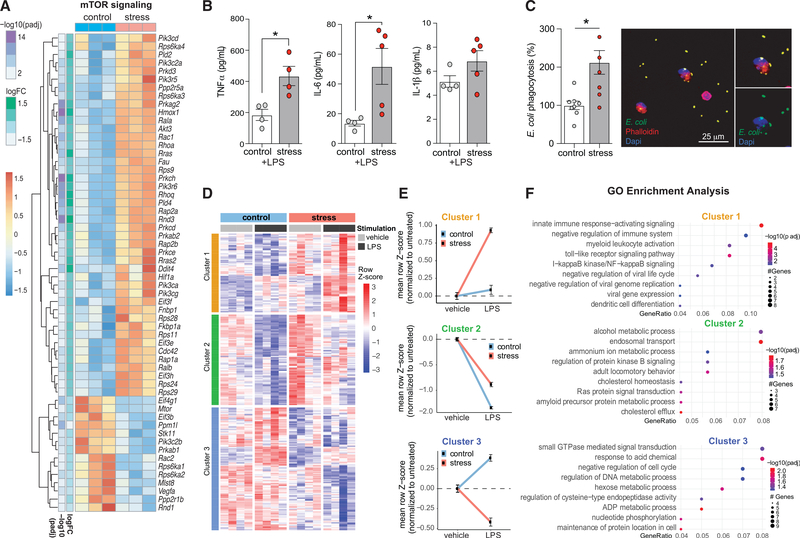
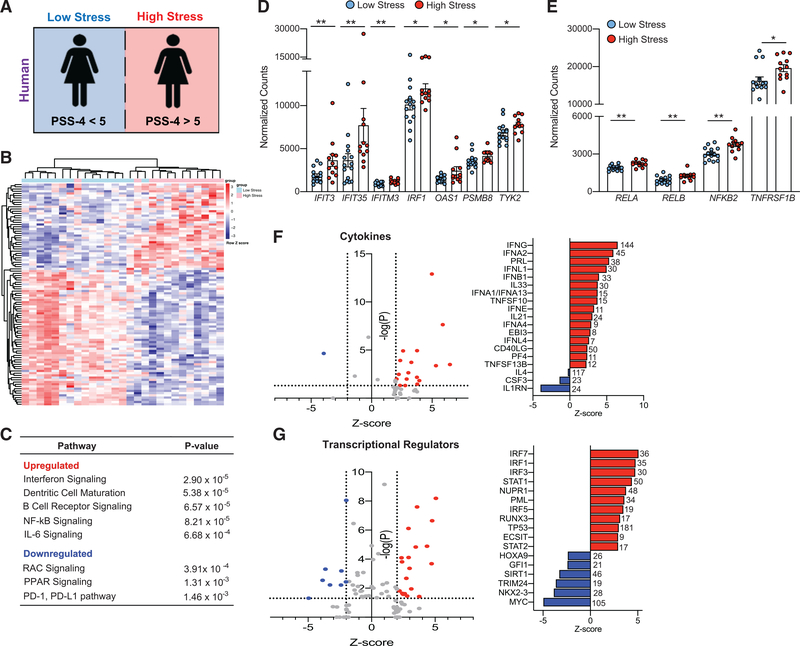
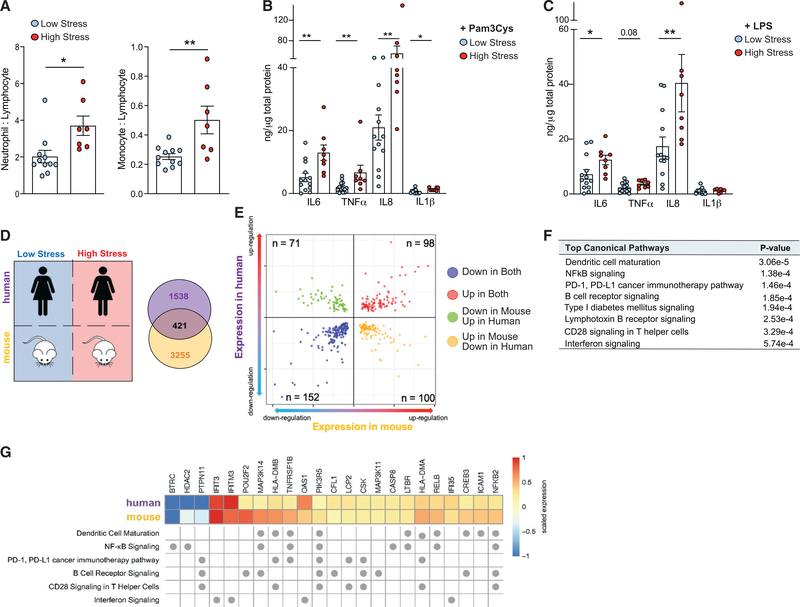
Psychological stress (PS) is associated with systemic inflammation and accelerates inflammatory disease progression (e.g., atherosclerosis). The mechanisms underlying stress-mediated inflammation and future health risk are poorly understood. Monocytes are key in sustaining systemic inflammation, and recent studies demonstrate that they maintain the memory of inflammatory insults, leading to a heightened inflammatory response upon rechallenge. We show that PS induces remodeling of the chromatin landscape and transcriptomic reprogramming of monocytes, skewing them to a primed hyperinflammatory phenotype. Monocytes from stressed mice and humans exhibit a characteristic inflammatory transcriptomic signature and are hyperresponsive upon stimulation with Toll-like receptor ligands. RNA and ATAC sequencing reveal that monocytes from stressed mice and humans exhibit activation of metabolic pathways (mTOR and PI3K) and reduced chromatin accessibility at mitochondrial respiration-associated loci. Collectively, our findings suggest that PS primes the reprogramming of myeloid cells to a hyperresponsive inflammatory state, which may explain how PS confers inflammatory disease risk.
Trial registration: ClinicalTrials.gov NCT03022552.
Keywords: inflammation; metabolism; monocytes; priming; psychological stress; women.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests M.N. has received funds or material research support from Lilly, Alnylam, Biotronik, CSL Behring, GlycoMimetics, GSK, Medtronic, Novartis, and Pfizer, as well as consulting fees from Biogen, Gimv, IFM Therapeutics, Molecular Imaging, Sigilon, and Verseau Therapeutics. H.R.R. receives in-kind donations for research from Abbott Vascular, Siemens, and BioTelemetry. All of the other authors declare no competing interests.
Figures
Comment in
-
Chronic stress and inflammation.Lab Anim (NY). 2021 Nov;50(11):309. doi: 10.1038/s41684-021-00884-y. Lab Anim (NY). 2021. PMID: 34697441 No abstract available.
References
-
- Ahmadi N, Hajsadeghi F, Mirshkarlo HB, Budoff M, Yehuda R, and Ebrahimi R (2011). Post-traumatic stress disorder, coronary atherosclerosis, and mortality. Am. J. Cardiol 108, 29–33. - PubMed
-
- Andrews S (2010). FastQC: a quality control tool for high throughput sequence data, Available online at. http://www.bioinformatics.babraham.ac.uk/projects/fastqc.
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
