

A simulation study of a honeybee breeding scheme accounting for polyandry, direct and maternal effects on colony performance
- PMID: 34496761
- PMCID: PMC8425095
- DOI: 10.1186/s12711-021-00665-8
A simulation study of a honeybee breeding scheme accounting for polyandry, direct and maternal effects on colony performance
Abstract
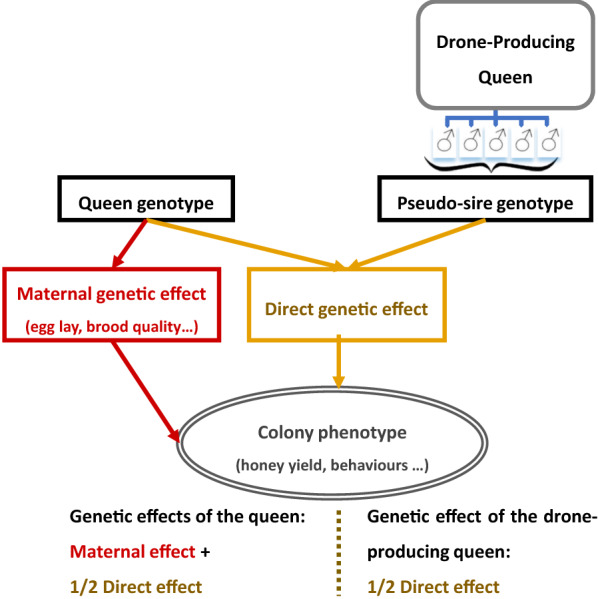
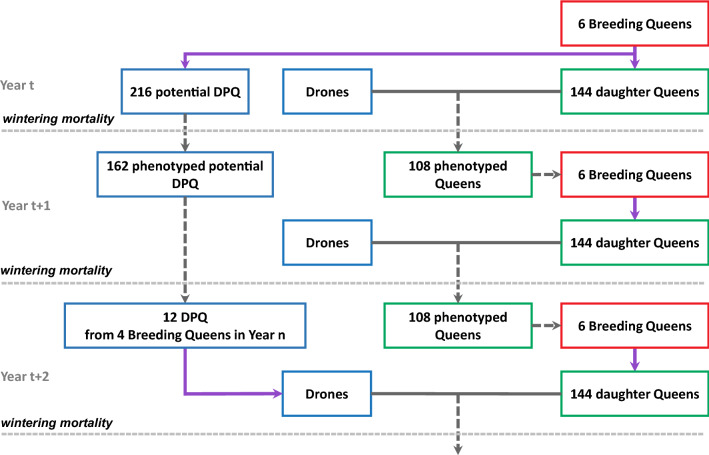
Background: Efficient breeding programs are difficult to implement in honeybees due to their biological specificities (polyandry and haplo-diploidy) and complexity of the traits of interest, with performances being measured at the colony scale and resulting from the joint effects of tens of thousands of workers (called direct effects) and of the queen (called maternal effects). We implemented a Monte Carlo simulation program of a breeding plan designed specifically for Apis mellifera's populations to assess the impact of polyandry versus monoandry on colony performance, inbreeding level and genetic gain depending on the individual selection strategy considered, i.e. complete mass selection or within-family (maternal lines) selection. We simulated several scenarios with different parameter setups by varying initial genetic variances and correlations between direct and maternal effects, the selection strategy and the polyandry level. Selection was performed on colony phenotypes.
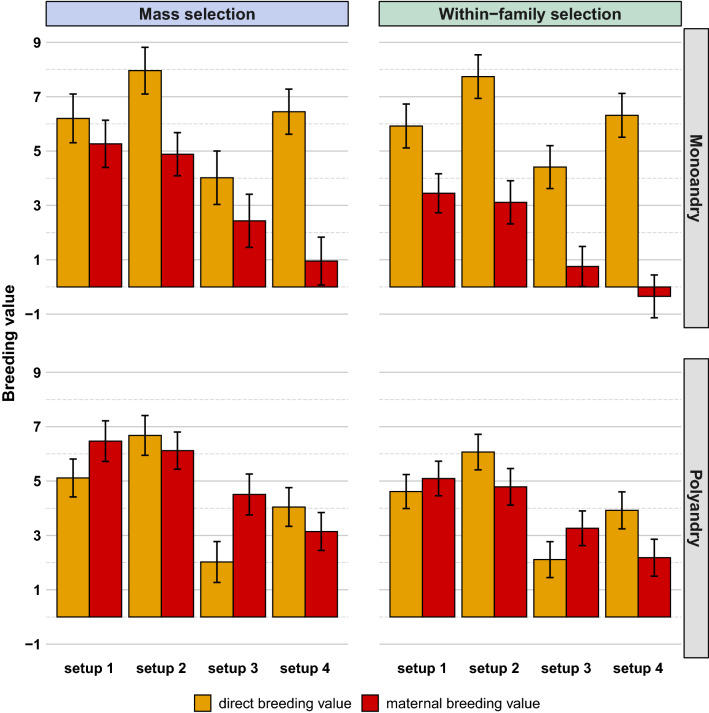
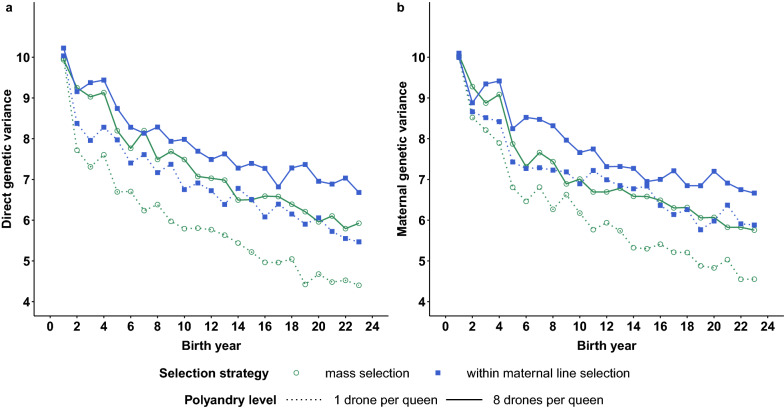
Results: All scenarios showed strong increases in direct breeding values of queens after 20 years of selection. Monoandry led to significantly higher direct than maternal genetic gains, especially when a negative correlation between direct and maternal effects was simulated. However, the relative increase in these genetic gains depended also on their initial genetic variability and on the selection strategy. When polyandry was simulated, the results were very similar with either 8 or 16 drones mated to each queen. Across scenarios, polyandrous mating resulted in equivalent or higher gains in performance than monoandrous mating, but with considerably lower inbreeding rates. Mass selection conferred a ~ 20% increase in performance compared to within-family selection, but was also accompanied by a strong increase in inbreeding levels (25 to 50% higher).
Conclusions: Our study is the first to compare the long-term effects of polyandrous versus monoandrous mating in honeybee breeding. The latter is an emergent strategy to improve specific traits, such as resistance to varroa, which can be difficult or expensive to phenotype. However, if used during several generations in a closed population, monoandrous mating increases the inbreeding level of queens much more than polyandrous mating, which is a strong limitation of this strategy.
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
MeSH terms
LinkOut - more resources
Full Text Sources
