

Metabolic flexibility during sleep
- PMID: 34497320
- PMCID: PMC8426397
- DOI: 10.1038/s41598-021-97301-8
Metabolic flexibility during sleep
Abstract
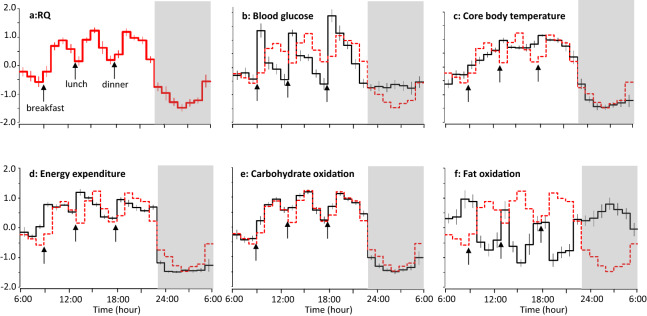
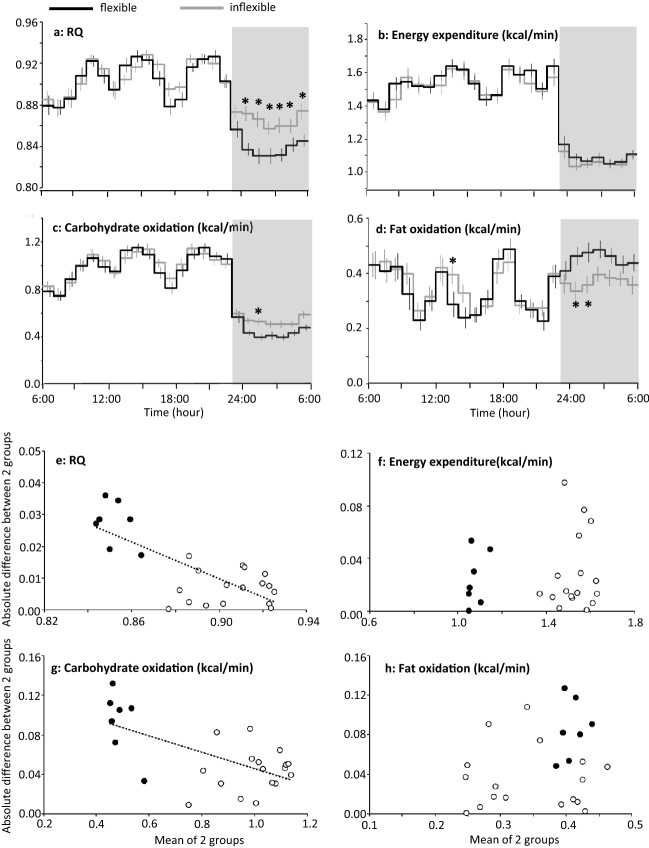
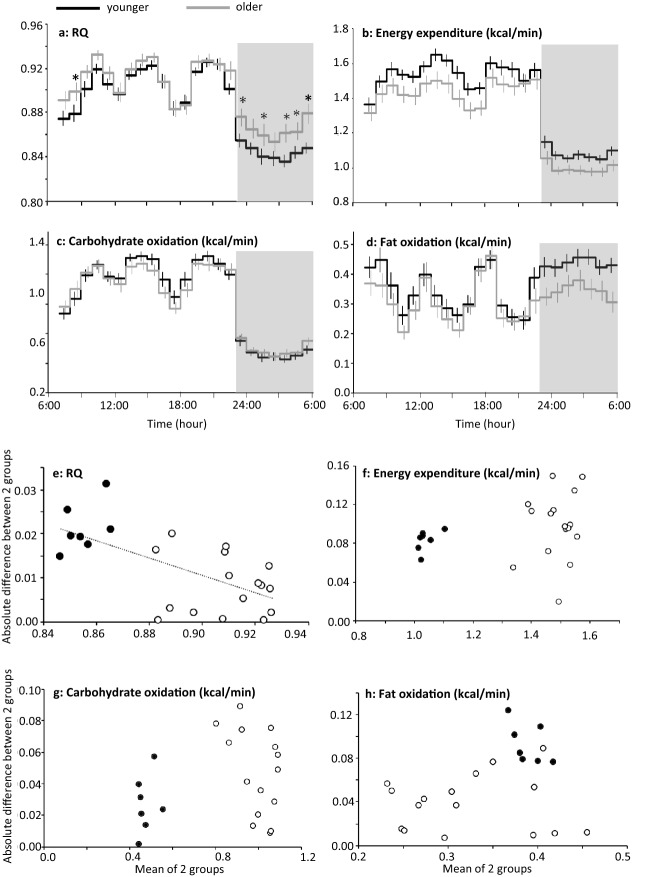
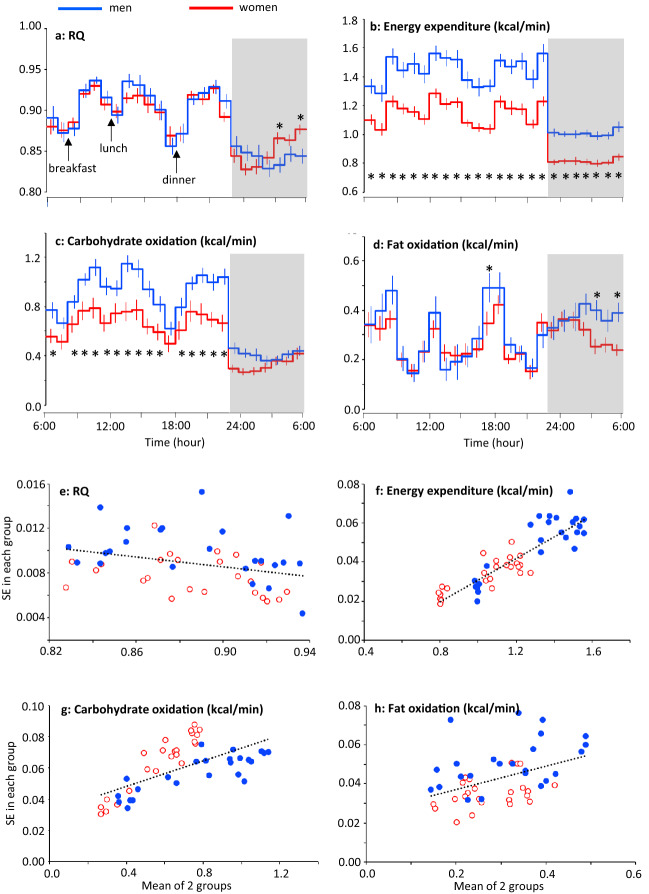
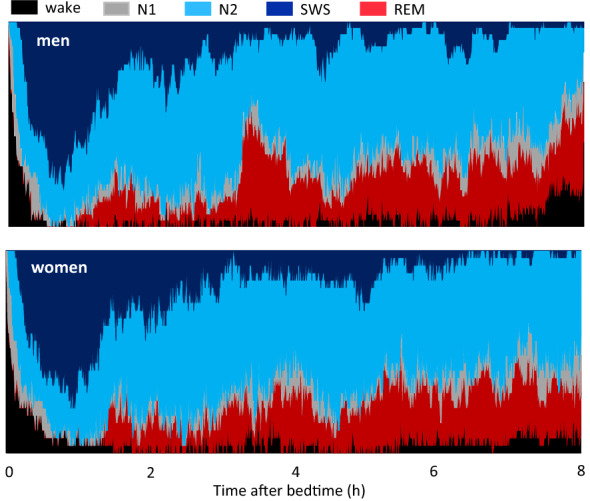
Known as metabolic flexibility, oxidized substrate is selected in response to changes in the nutritional state. Sleep imposes an extended duration of fasting, and oxidized substrates during sleep were assumed to progressively shift from carbohydrate to fat, thereby gradually decreasing the respiratory quotient (RQ). Contrary to this assumption, whole-room indirect calorimetry with improved time resolution revealed that RQ re-ascended prior to awakening, and nadir of RQ in non-obese young adults occurred earlier in women than men after bedtime. The transient decrease in RQ during sleep was blunted in metabolically inflexible men with smaller amplitude of diurnal rhythm in RQ. Similarly, the effect of 10 years difference in age on RQ became significant during sleep; the decrease in RQ during sleep was blunted in older subjects. Inter-individual difference in RQ become apparent during sleep, and it might serve as a window to gain insight into the early-stage pathogenesis of metabolic inflexibility.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
