

MAIT Cells Balance the Requirements for Immune Tolerance and Anti-Microbial Defense During Pregnancy
- PMID: 34497611
- PMCID: PMC8420809
- DOI: 10.3389/fimmu.2021.718168
MAIT Cells Balance the Requirements for Immune Tolerance and Anti-Microbial Defense During Pregnancy
Abstract
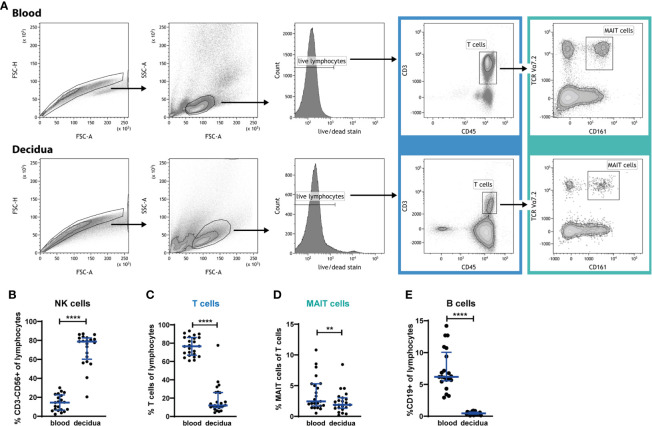
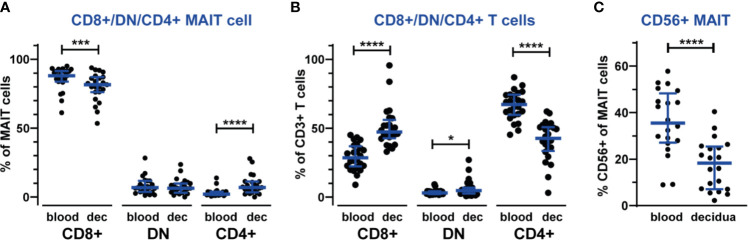
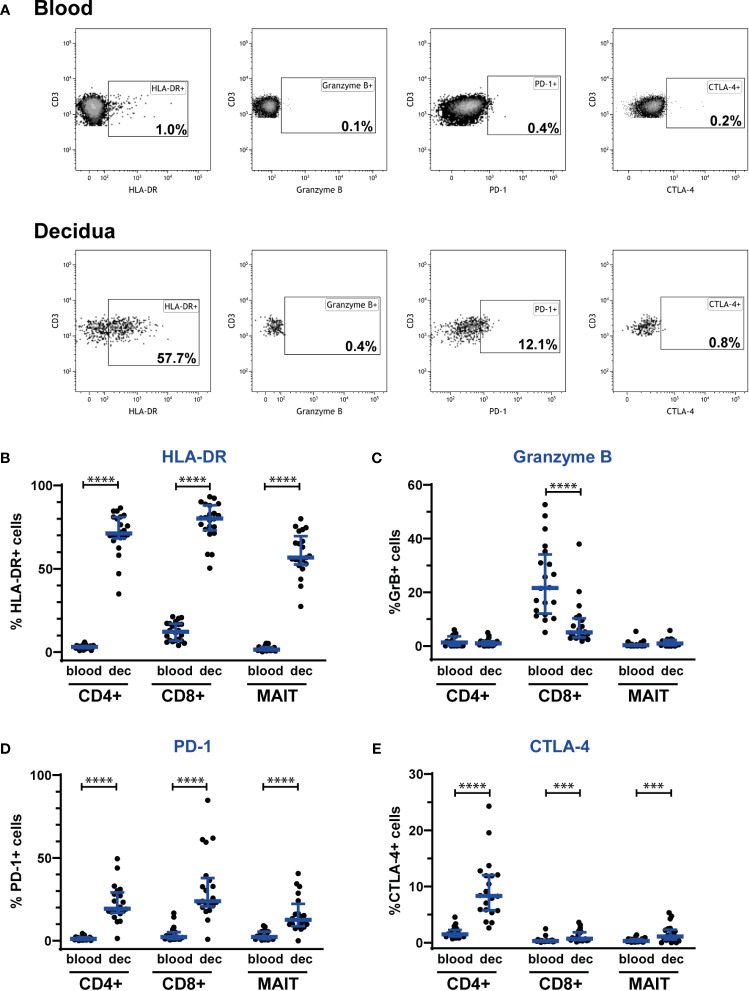
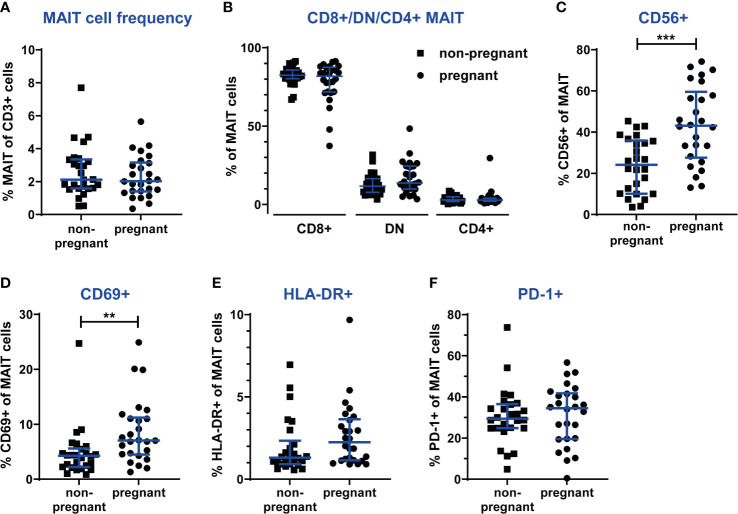
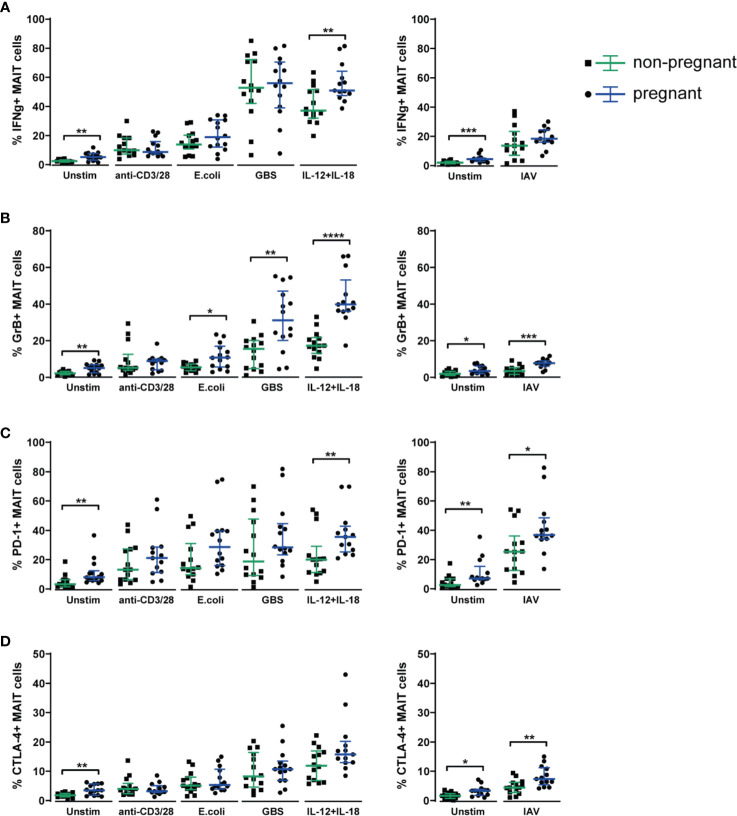
Mucosal-associated invariant T (MAIT) cells are an innate-like T cell subset with proinflammatory and cytotoxic effector functions. During pregnancy, modulation of the maternal immune system, both at the fetal-maternal interface and systemically, is crucial for a successful outcome and manifests through controlled enhancement of innate and dampening of adaptive responses. Still, immune defenses need to efficiently protect both the mother and the fetus from infection. So far, it is unknown whether MAIT cells are subjected to immunomodulation during pregnancy, and characterization of decidual MAIT cells as well as their functional responses during pregnancy are mainly lacking. We here characterized the presence and phenotype of Vα7.2+CD161+ MAIT cells in blood and decidua (the uterine endometrium during pregnancy) from women pregnant in the 1st trimester, i.e., the time point when local immune tolerance develops. We also assessed the phenotype and functional responses of MAIT cells in blood of women pregnant in the 3rd trimester, i.e., when systemic immunomodulation is most pronounced. Multi-color flow cytometry panels included markers for MAIT subsets, and markers of activation (CD69, HLA-DR, Granzyme B) and immunoregulation (PD-1, CTLA-4). MAIT cells were numerically decreased at the fetal-maternal interface and showed, similar to other T cells in the decidua, increased expression of immune checkpoint markers compared with MAIT cells in blood. During the 3rd trimester, circulating MAIT cells showed a higher expression of CD69 and CD56, and their functional responses to inflammatory (activating anti-CD3/CD28 antibodies, and IL-12 and IL-18) and microbial stimuli (Escherichia coli, group B streptococci and influenza A virus) were generally increased compared with MAIT cells from non-pregnant women, indicating enhanced antimicrobial defenses during pregnancy. Taken together, our findings indicate dual roles for MAIT cells during pregnancy, with an evidently well-adapted ability to balance the requirements of immune tolerance in parallel with maintained antimicrobial defenses. Since MAIT cells are easily activated, they need to be strictly regulated during pregnancy, and failure to do so could contribute to pregnancy complications.
Keywords: T cells; anti-microbial response; decidua; immune tolerance; immunomodulation; innate-like T cells; mucosal-associated invariant T (MAIT) cells; pregnancy.
Copyright © 2021 Raffetseder, Lindau, van der Veen, Berg, Larsson and Ernerudh.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
MAIT cells accumulate in placental intervillous space and display a highly cytotoxic phenotype upon bacterial stimulation.Sci Rep. 2017 Jul 21;7(1):6123. doi: 10.1038/s41598-017-06430-6. Sci Rep. 2017. PMID: 28733576 Free PMC article.
-
Human MAIT Cell Activation In Vitro.Methods Mol Biol. 2020;2098:97-124. doi: 10.1007/978-1-0716-0207-2_7. Methods Mol Biol. 2020. PMID: 31792818
-
Immune Profile of the Normal Maternal-Fetal Interface in Rhesus Macaques and Its Alteration Following Zika Virus Infection.Front Immunol. 2021 Jul 29;12:719810. doi: 10.3389/fimmu.2021.719810. eCollection 2021. Front Immunol. 2021. PMID: 34394129 Free PMC article.
-
MAIT Cells at the Fetal-Maternal Interface During Pregnancy.Front Immunol. 2020 Aug 19;11:1788. doi: 10.3389/fimmu.2020.01788. eCollection 2020. Front Immunol. 2020. PMID: 32973750 Free PMC article. Review.
-
The Immune Modulating Properties of Mucosal-Associated Invariant T Cells.Front Immunol. 2020 Aug 13;11:1556. doi: 10.3389/fimmu.2020.01556. eCollection 2020. Front Immunol. 2020. PMID: 32903532 Free PMC article. Review.
Cited by
-
Labour promotes systemic mobilisation of monocytes, T cell activation and local secretion of chemotactic factors in the intervillous space of the placenta.Front Immunol. 2023 Mar 8;14:1129261. doi: 10.3389/fimmu.2023.1129261. eCollection 2023. Front Immunol. 2023. PMID: 36969250 Free PMC article.
-
Single-cell RNA-sequencing highlights a curtailed NK cell function in convalescent COVID-19 pregnant women.Front Immunol. 2025 Jun 30;16:1560391. doi: 10.3389/fimmu.2025.1560391. eCollection 2025. Front Immunol. 2025. PMID: 40661959 Free PMC article.
-
Altered Cytokine Production in Human Intervillous Blood T Cells in Preeclampsia.Reprod Sci. 2023 Sep;30(9):2655-2664. doi: 10.1007/s43032-023-01165-4. Epub 2023 Feb 7. Reprod Sci. 2023. PMID: 36749459 Free PMC article.
-
The dose- and time-dependent effectiveness and safety associated with COVID-19 vaccination during pregnancy: a systematic review and meta-analysis.Int J Infect Dis. 2023 Mar;128:335-346. doi: 10.1016/j.ijid.2023.01.018. Epub 2023 Jan 25. Int J Infect Dis. 2023. PMID: 36707044 Free PMC article.
-
Immune profiling of SARS-CoV-2 infection during pregnancy reveals NK cell and γδ T cell perturbations.JCI Insight. 2023 Apr 10;8(7):e167157. doi: 10.1172/jci.insight.167157. JCI Insight. 2023. PMID: 37036008 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous