

Challenges on the development of a pseudotyping assay for Zika glycoproteins
- PMID: 34499027
- PMCID: PMC8697511
- DOI: 10.1099/jmm.0.001413
Challenges on the development of a pseudotyping assay for Zika glycoproteins
Abstract
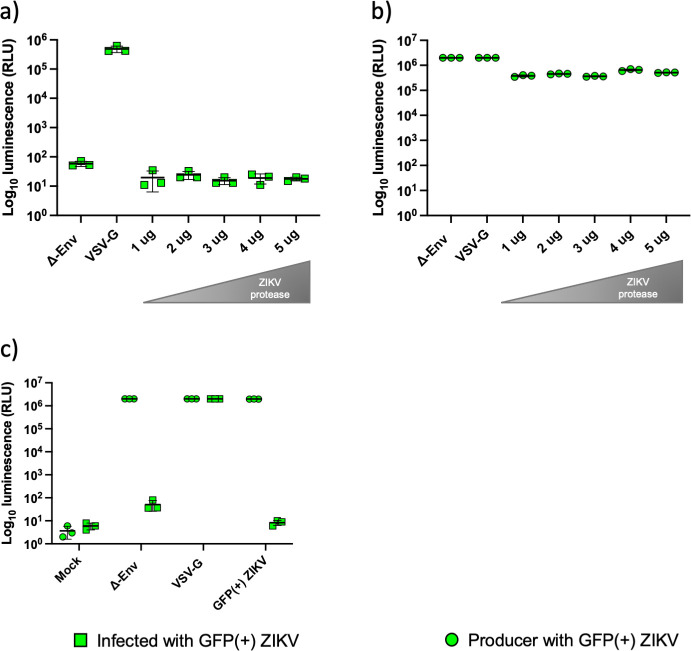
Introduction. Zika virus (ZIKV) emerged as a public health concern on the American continent during late 2015. As the number of infected grew so did the concerns about its capability to cause long-term damage especially with the appearance of the congenital Zika syndrome (CZS). Proteins from the TAM family of receptor tyrosine kinases (RTKs) were proposed as the cellular receptors, however, due to the ability of the virus to infect a variety of cell lines different strategies to elucidate the tropism of the virus should be investigated.Hypothesis. Pseudotyping is a powerful tool to interrogate the ability of the glycoprotein (GP) to permit entry of viruses.Aim. We aimed to establish a highly tractable pseudotype model using lenti- and retro-viral backbones to investigate the entry pathway of ZIKV.Methodology. We used different glycoprotein constructs and different lenti- or retro-viral backbones, in a matrix of ratios to investigate production of proteins and functional pseudotypes.Results. Varying the ratio of backbone and glycoprotein plasmids did not yield infectious pseudotypes. Moreover, the supplementation of the ZIKV protease or the substitution of the backbone had no positive impact on the infectivity. We showed production of the proteins in producer cells implying the lack of infectious pseudotypes is due to a lack of successful glycoprotein incorporation, rather than lack of protein production.Conclusion. In line with other reports, we were unable to successfully produce infectious pseudotypes using the variety of methods described. Other strategies may be more suitable in the development of an efficient pseudotype model for ZIKV and other flaviviruses.
Keywords: Zika virus; glycoprotein; pseudotype.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures
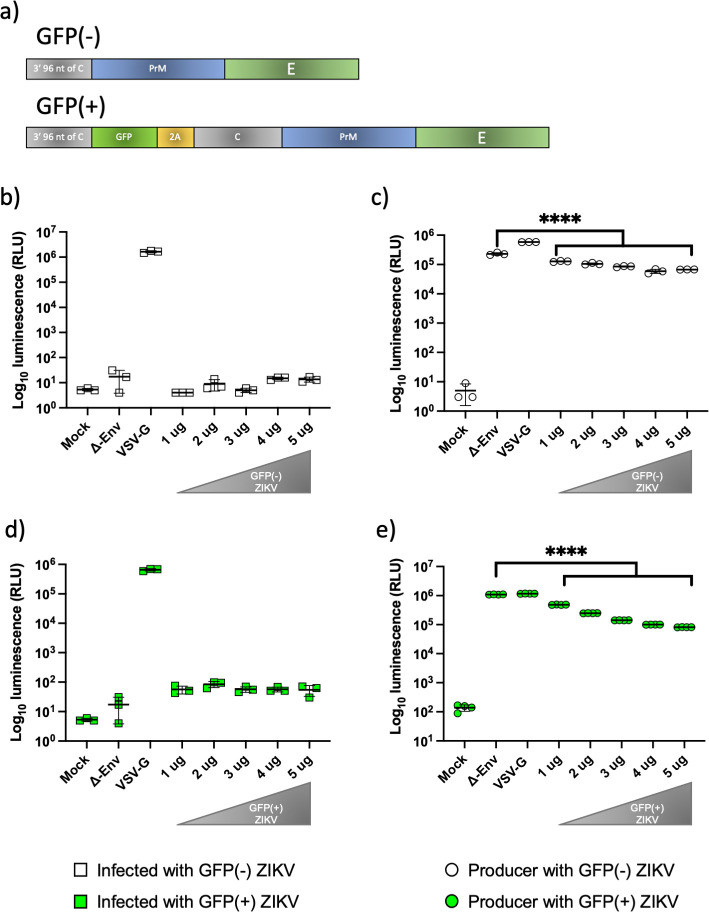
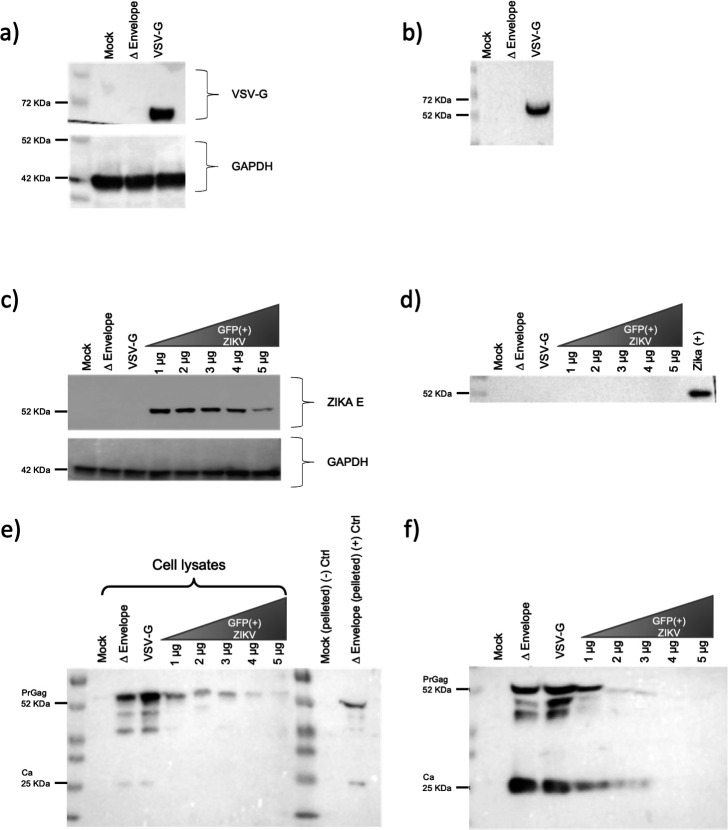
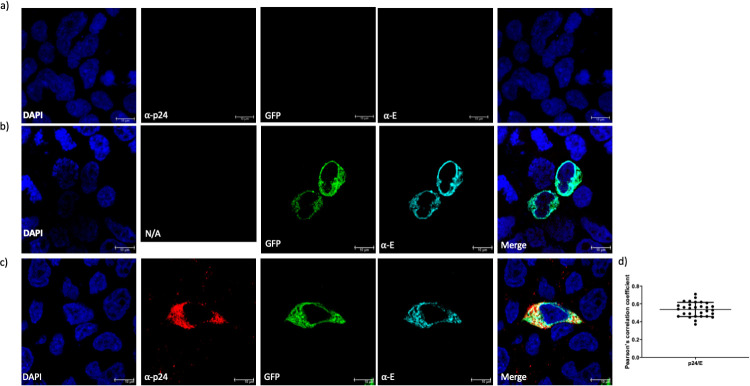
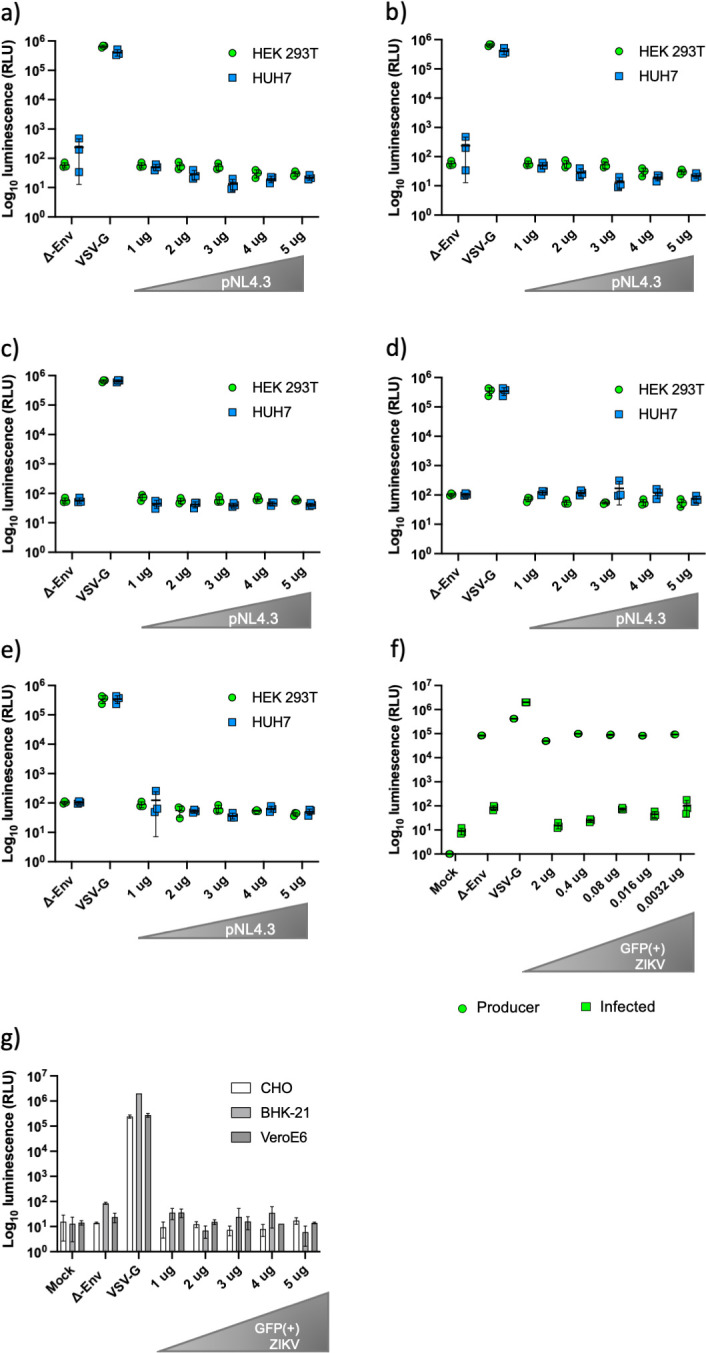
References
-
- Gebre Y, Forbes N, Gebre T. Zika virus infection, transmission, associated neurological disorders and birth abnormalities: A review of progress in research, priorities and knowledge gaps. Asian Pac J Trop Biomed. 2016;6:815–824. doi: 10.1016/j.apjtb.2016.08.008. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials