

Low lysophosphatidylcholine induces skeletal muscle myopathy that is aggravated by high-fat diet feeding
- PMID: 34499764
- PMCID: PMC8439548
- DOI: 10.1096/fj.202101104R
Low lysophosphatidylcholine induces skeletal muscle myopathy that is aggravated by high-fat diet feeding
Abstract
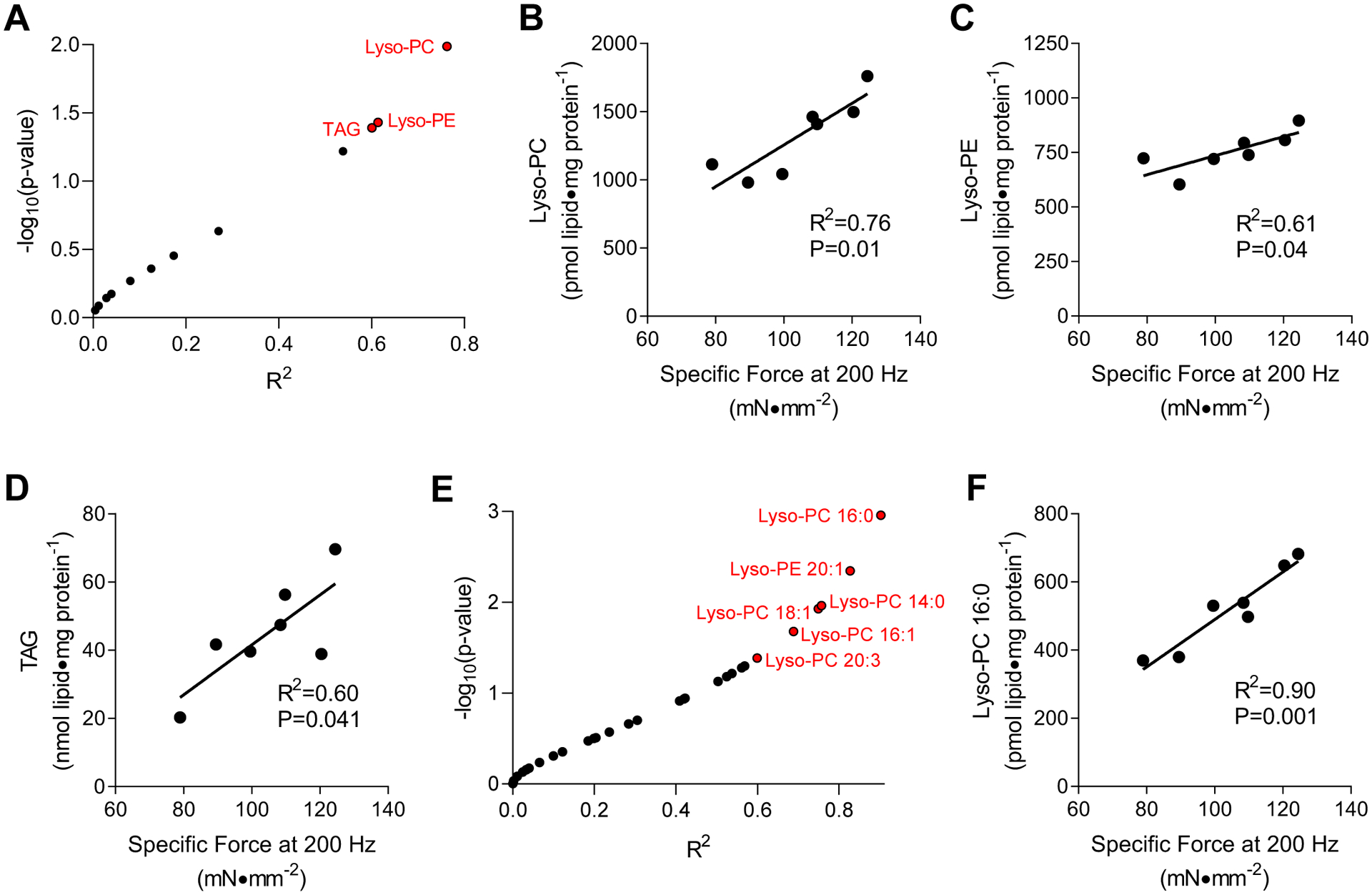
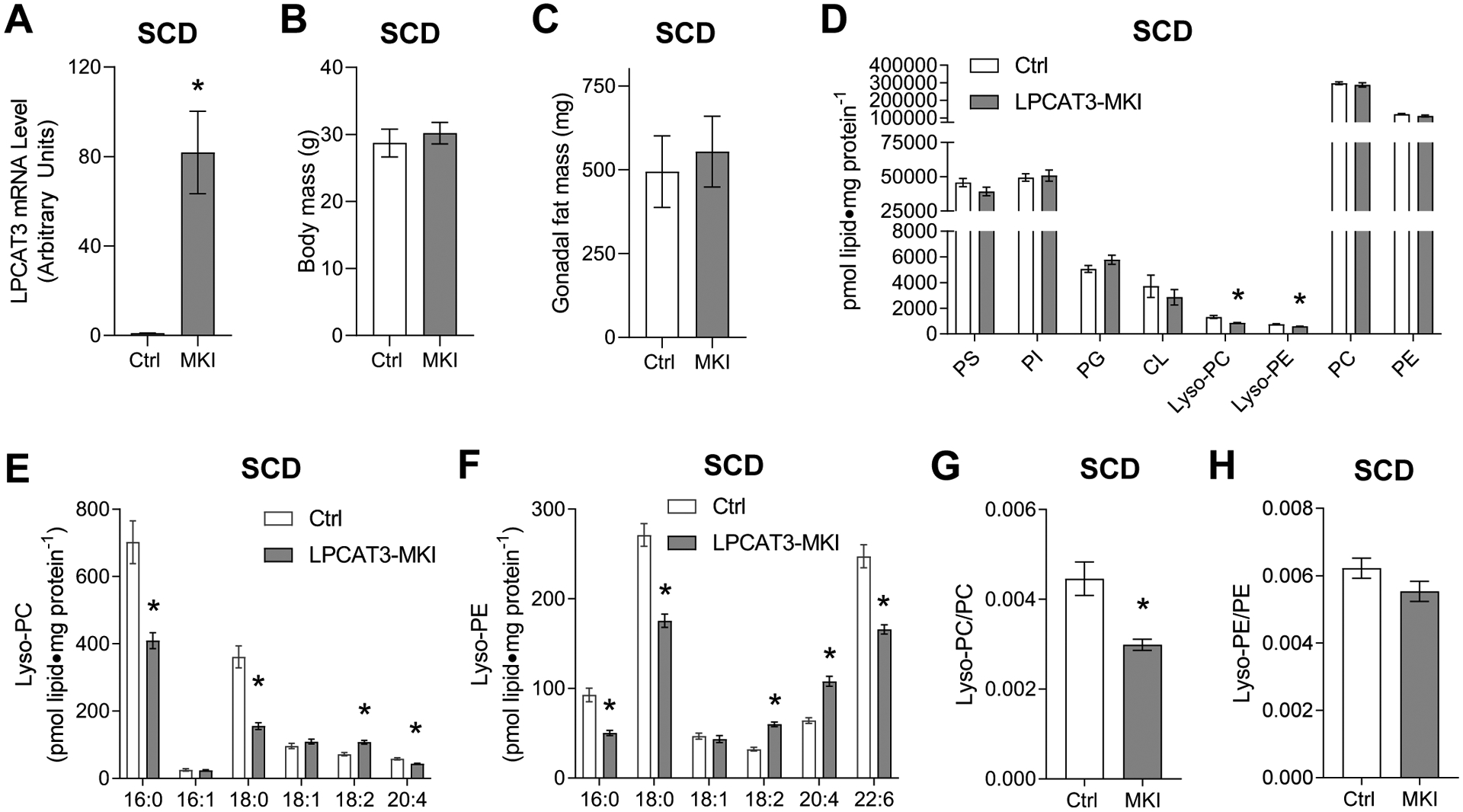
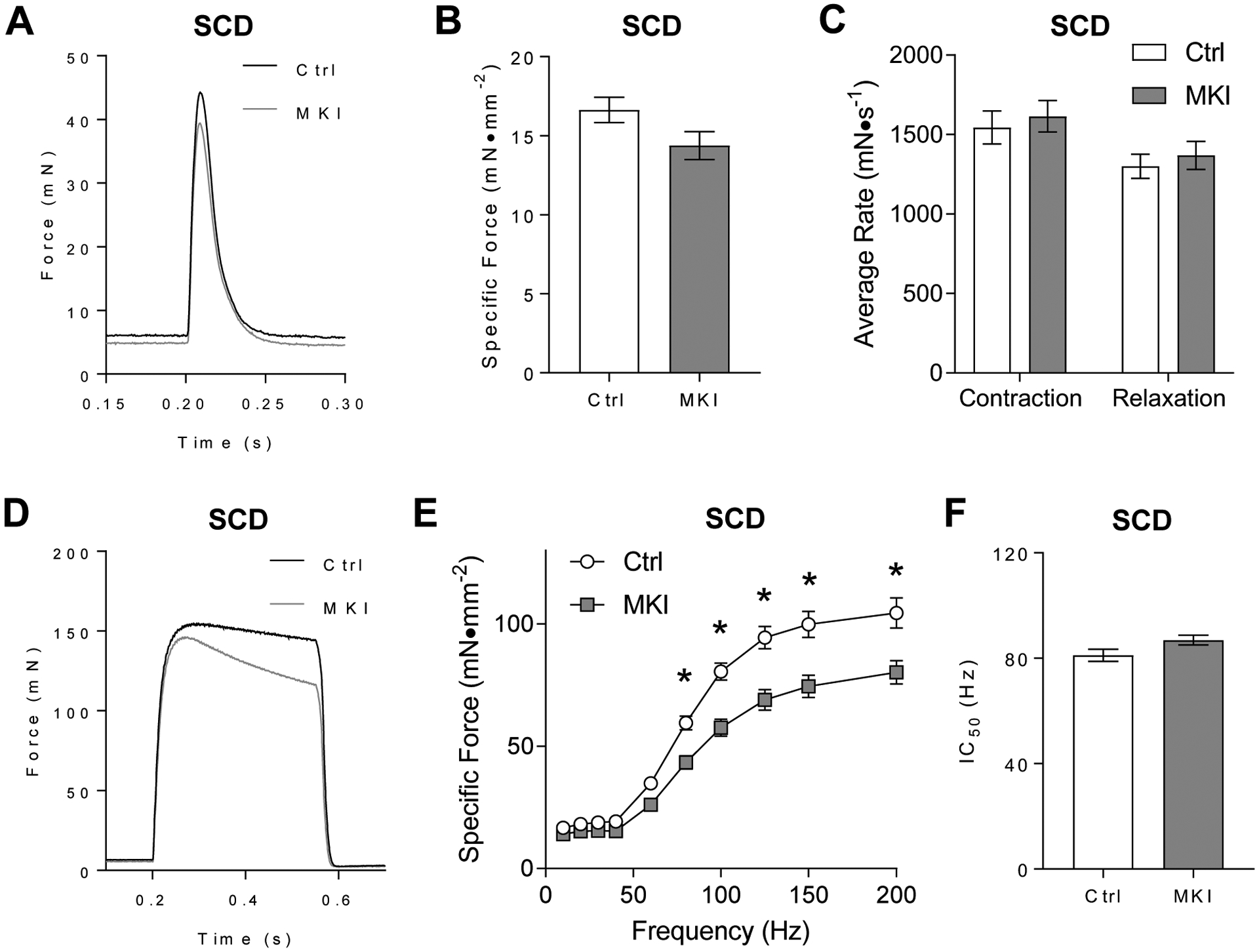
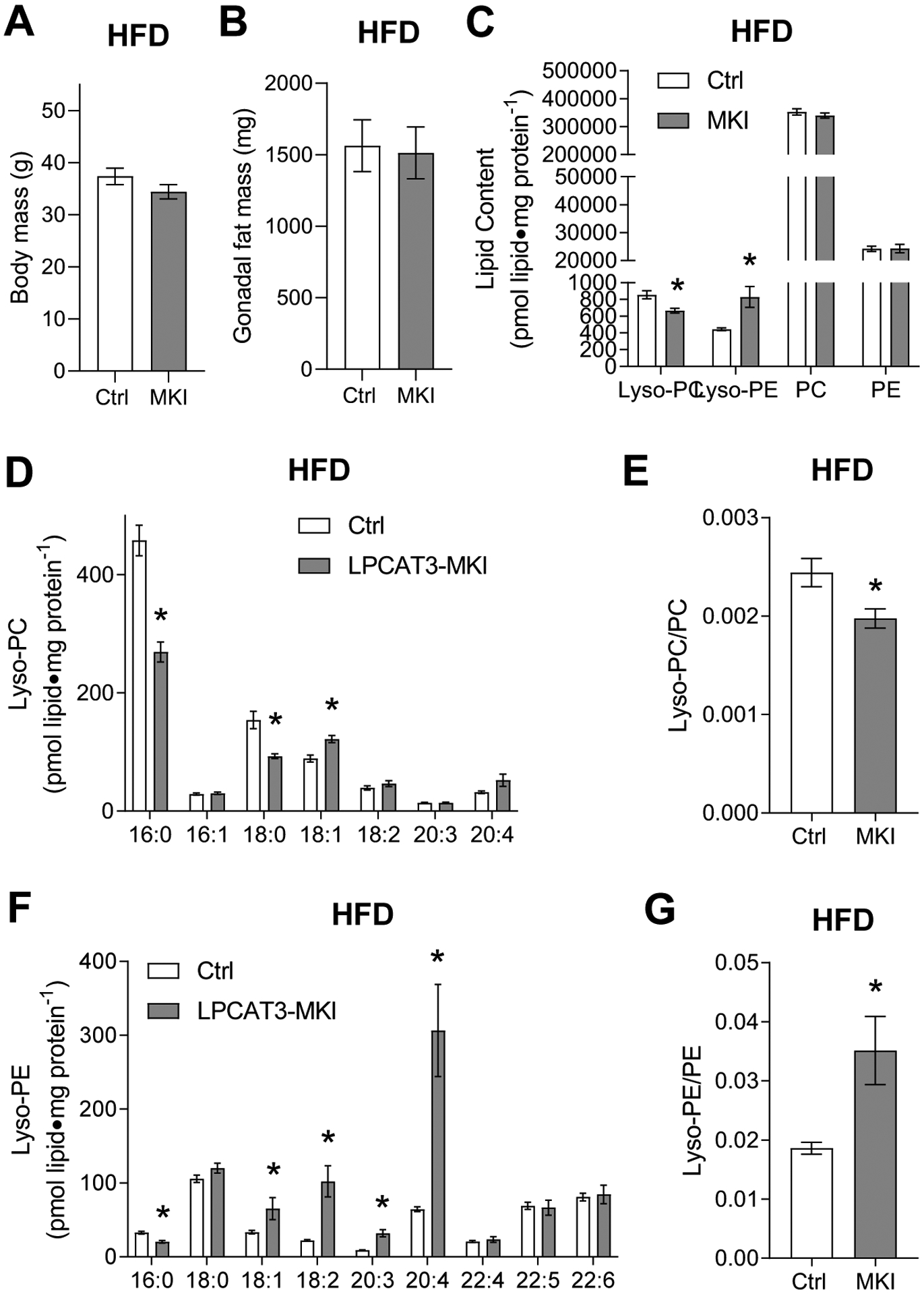
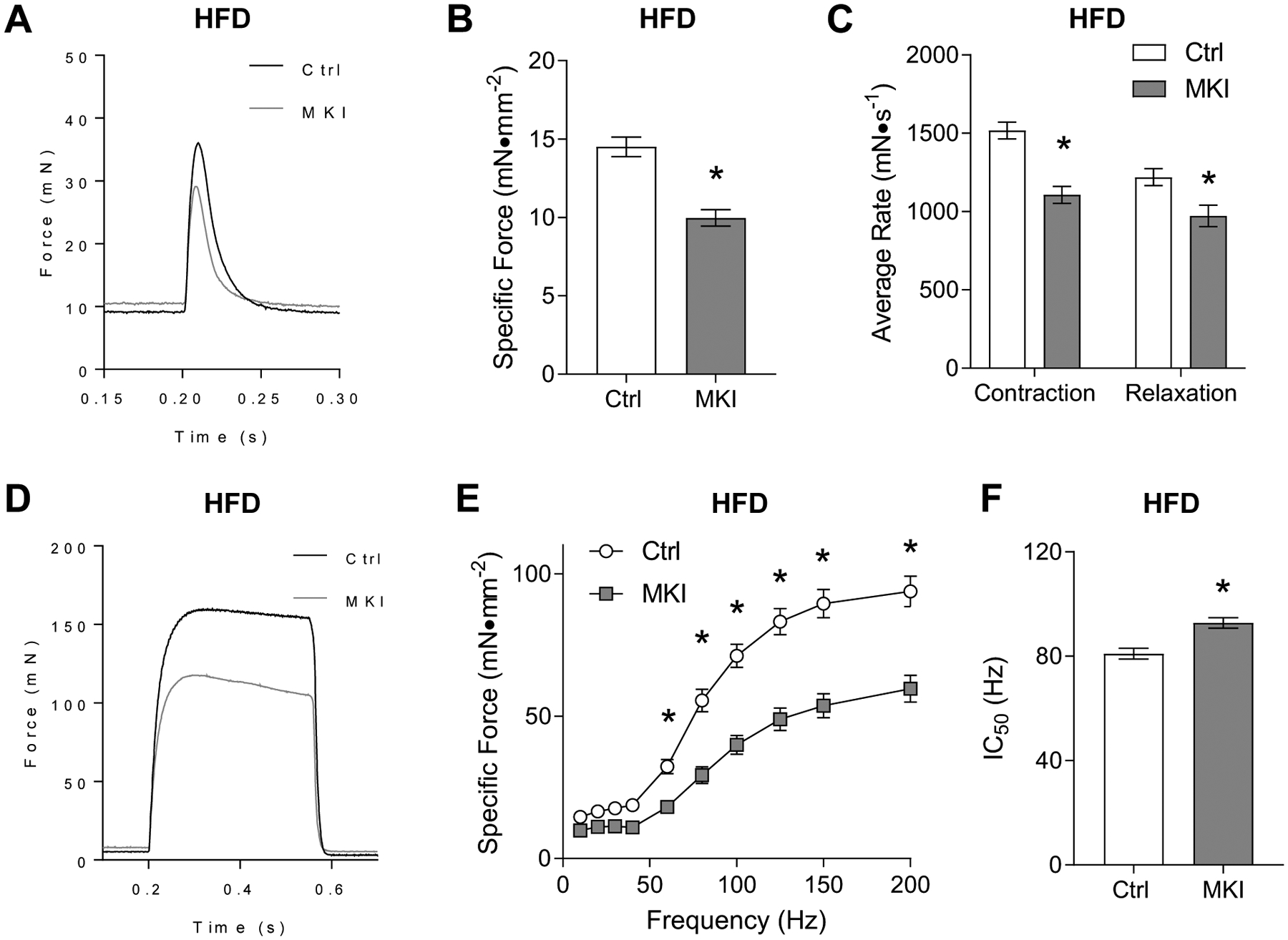
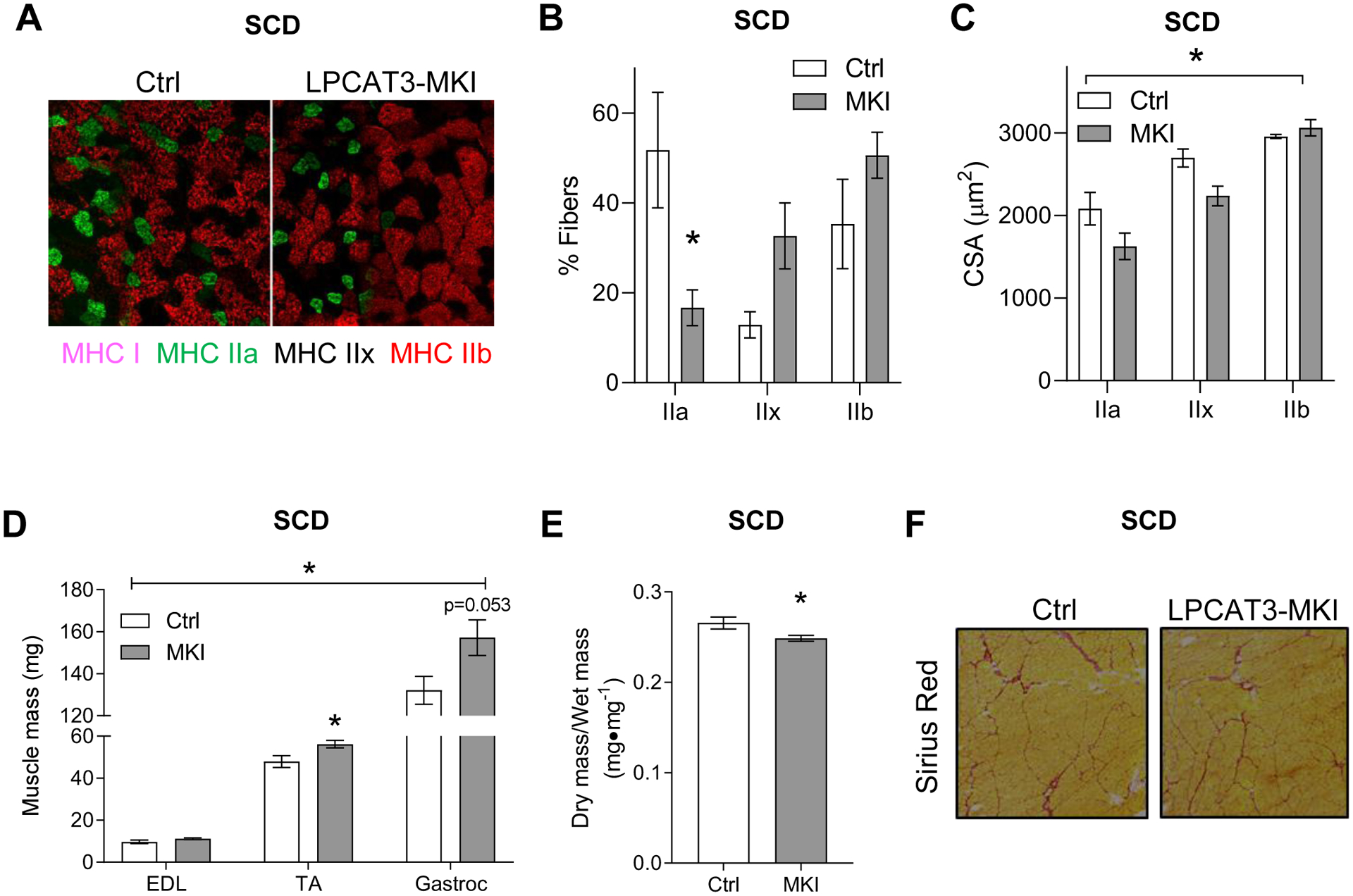
Obesity alters skeletal muscle lipidome and promotes myopathy, but it is unknown whether aberrant muscle lipidome contributes to the reduction in skeletal muscle contractile force-generating capacity. Comprehensive lipidomic analyses of mouse skeletal muscle revealed a very strong positive correlation between the abundance of lysophosphatidylcholine (lyso-PC), a class of lipids that is known to be downregulated with obesity, with maximal tetanic force production. The level of lyso-PC is regulated primarily by lyso-PC acyltransferase 3 (LPCAT3), which acylates lyso-PC to form phosphatidylcholine. Tamoxifen-inducible skeletal muscle-specific overexpression of LPCAT3 (LPCAT3-MKI) was sufficient to reduce muscle lyso-PC content in both standard chow diet- and high-fat diet (HFD)-fed conditions. Strikingly, the assessment of skeletal muscle force-generating capacity ex vivo revealed that muscles from LPCAT3-MKI mice were weaker regardless of diet. Defects in force production were more apparent in HFD-fed condition, where tetanic force production was 40% lower in muscles from LPCAT3-MKI compared to that of control mice. These observations were partly explained by reductions in the cross-sectional area in type IIa and IIx fibers, and signs of muscle edema in the absence of fibrosis. Future studies will pursue the mechanism by which LPCAT3 may alter protein turnover to promote myopathy.
Keywords: diabetes; lysophospholipid; myopathy; skeletal muscle.
© 2021 Federation of American Societies for Experimental Biology.
Conflict of interest statement
Figures
References
-
- Powers SK, Kavazis AN, and DeRuisseau KC (2005) Mechanisms of disuse muscle atrophy: role of oxidative stress. Am J Physiol Regul Integr Comp Physiol 288, R337–344 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R56 AG050781/AG/NIA NIH HHS/United States
- S10 OD016232/OD/NIH HHS/United States
- R03 DK109888/DK/NIDDK NIH HHS/United States
- R01 GM144613/GM/NIGMS NIH HHS/United States
- R01 AG050781/AG/NIA NIH HHS/United States
- R01 DK127979/DK/NIDDK NIH HHS/United States
- R21 AG063077/AG/NIA NIH HHS/United States
- P30 DK020579/DK/NIDDK NIH HHS/United States
- S10 OD021505/OD/NIH HHS/United States
- R01 AG074535/AG/NIA NIH HHS/United States
- R01 HL125695/HL/NHLBI NIH HHS/United States
- R01 DK107397/DK/NIDDK NIH HHS/United States
- U54 DK110858/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
