

Mitochondrial bioenergetics boost macrophage activation, promoting liver regeneration in metabolically compromised animals
- PMID: 34510498
- PMCID: PMC9300136
- DOI: 10.1002/hep.32149
Mitochondrial bioenergetics boost macrophage activation, promoting liver regeneration in metabolically compromised animals
Erratum in
-
Erratum.Hepatology. 2022 Aug;76(2):532. doi: 10.1002/hep.32505. Epub 2022 Apr 22. Hepatology. 2022. PMID: 35451511 Free PMC article. No abstract available.
Abstract
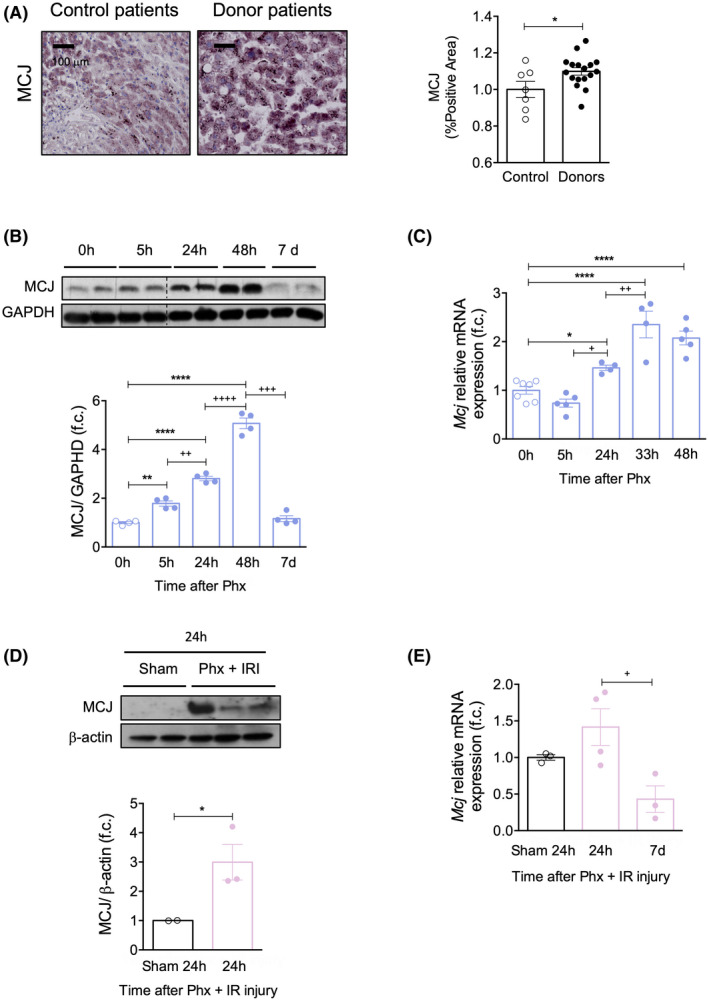
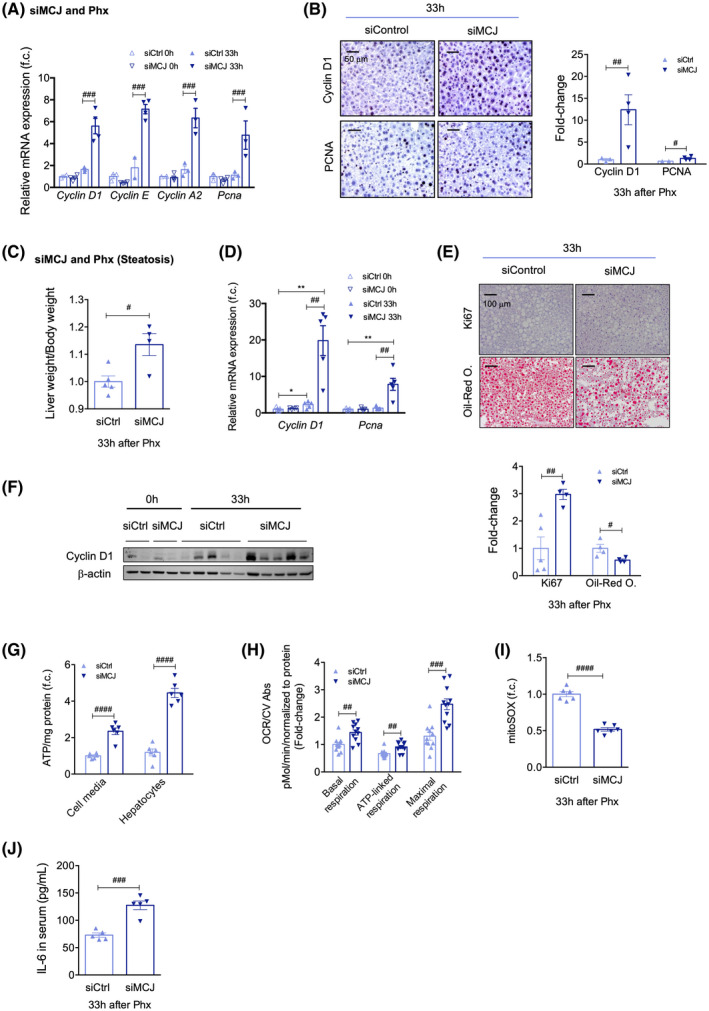
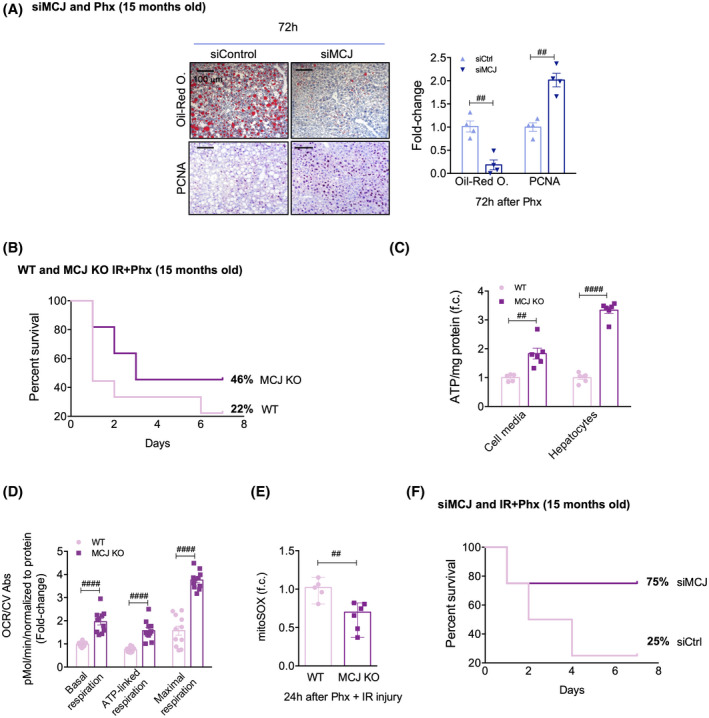
Background and aims: Hepatic ischemia-reperfusion injury (IRI) is the leading cause of early posttransplantation organ failure as mitochondrial respiration and ATP production are affected. A shortage of donors has extended liver donor criteria, including aged or steatotic livers, which are more susceptible to IRI. Given the lack of an effective treatment and the extensive transplantation waitlist, we aimed at characterizing the effects of an accelerated mitochondrial activity by silencing methylation-controlled J protein (MCJ) in three preclinical models of IRI and liver regeneration, focusing on metabolically compromised animal models.
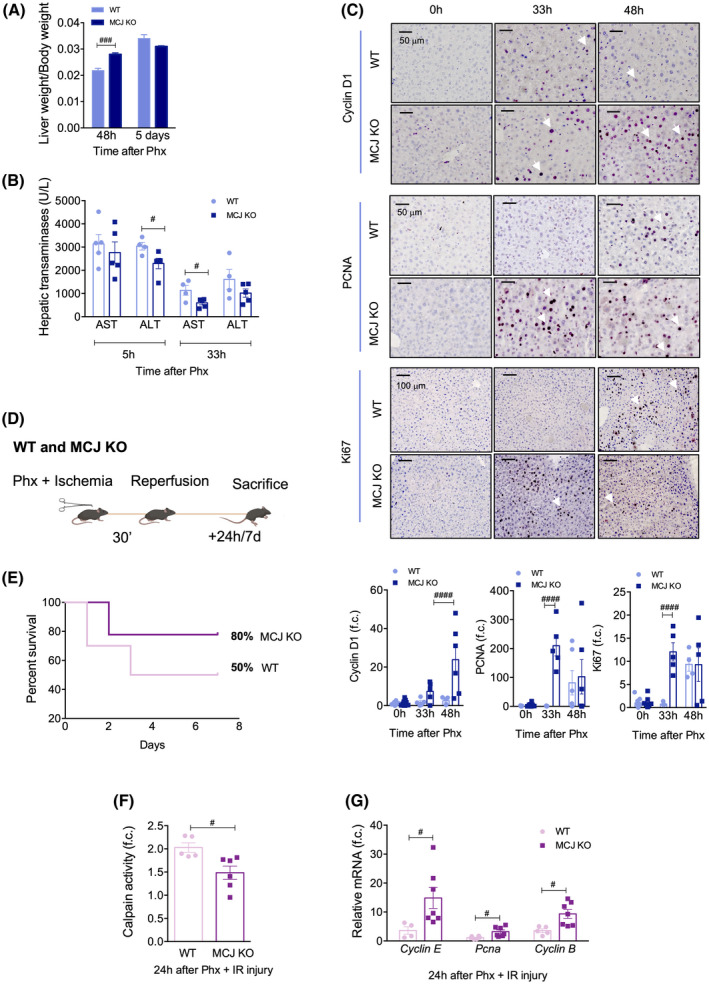
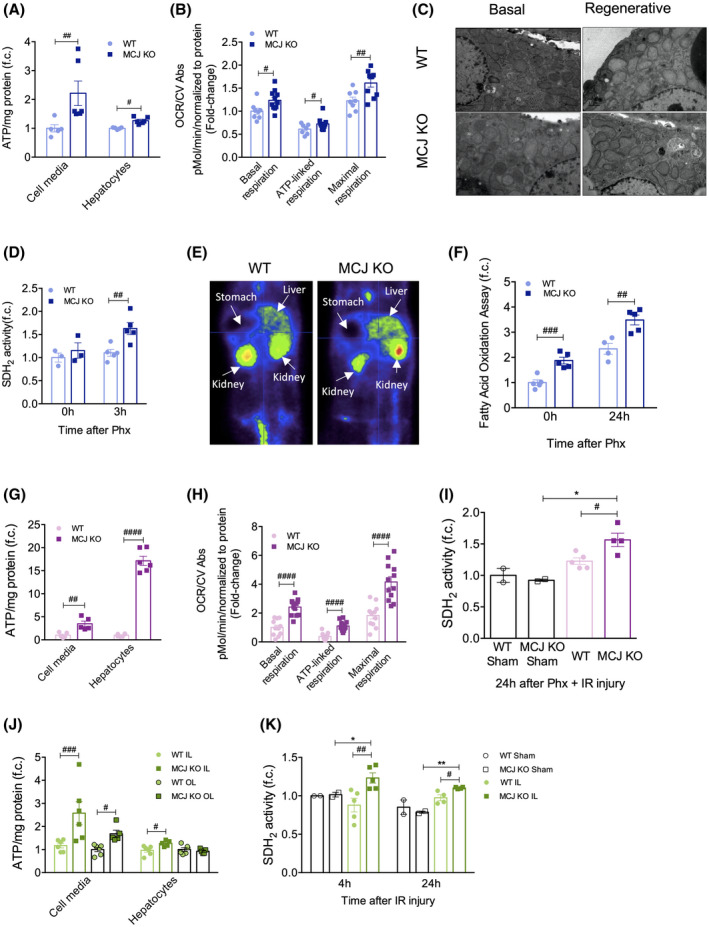
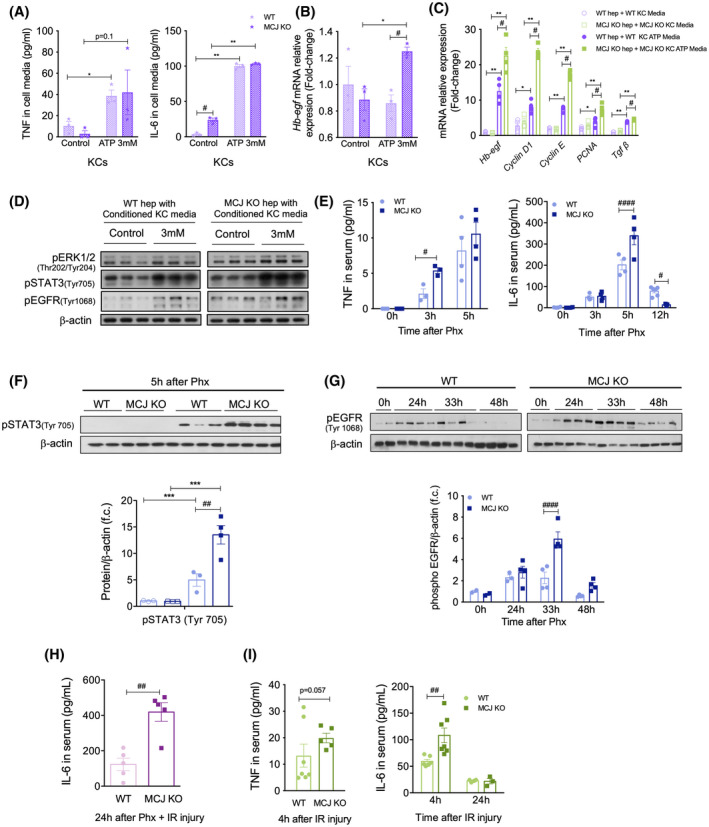
Approach and results: Wild-type (WT), MCJ knockout (KO), and Mcj silenced WT mice were subjected to 70% partial hepatectomy (Phx), prolonged IRI, and 70% Phx with IRI. Old and young mice with metabolic syndrome were also subjected to these procedures. Expression of MCJ, an endogenous negative regulator of mitochondrial respiration, increases in preclinical models of Phx with or without vascular occlusion and in donor livers. Mice lacking MCJ initiate liver regeneration 12 h faster than WT and show reduced ischemic injury and increased survival. MCJ knockdown enables a mitochondrial adaptation that restores the bioenergetic supply for enhanced regeneration and prevents cell death after IRI. Mechanistically, increased ATP secretion facilitates the early activation of Kupffer cells and production of TNF, IL-6, and heparin-binding EGF, accelerating the priming phase and the progression through G1 /S transition during liver regeneration. Therapeutic silencing of MCJ in 15-month-old mice and in mice fed a high-fat/high-fructose diet for 12 weeks improves mitochondrial respiration, reduces steatosis, and overcomes regenerative limitations.
Conclusions: Boosting mitochondrial activity by silencing MCJ could pave the way for a protective approach after major liver resection or IRI, especially in metabolically compromised, IRI-susceptible organs.
© 2021 The Authors. Hepatology published by Wiley Periodicals LLC on behalf of American Association for the Study of Liver Diseases.
Conflict of interest statement
Dr. Martínez‐Chantar advises for Mitotherapeutix LLC. All other authors have nothing to disclose.
Figures
References
-
- Trapero‐Marugán M, Little EC, Berenguer M. Stretching the boundaries for liver transplant in the 21st century. Lancet Gastroenterol Hepatol. 2018;3:803–11. - PubMed
-
- Younossi ZM, Koenig AB, Abdelatif D, Fazel Y, Henry L, Wymer M, et al. Global epidemiology of nonalcoholic fatty liver disease—meta‐analytic assessment of prevalence, incidende, and outcomes. Hepatology. 2016;64:73–84. - PubMed
-
- Vinaixa C, Selzner N, Berenguer M. Fat and liver transplantation: clinical implications. Transpl Int. 2018;31:828–37. - PubMed
-
- Jaime FD, Berenguer M, Fernando A, Martorell A. Pushing the donor limits: deceased donor liver transplantation using organs from octogenarian donors. Liver Transplant. 2017;23:S22–6. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
