

Regulation of biomolecular condensates by interfacial protein clusters
- PMID: 34516789
- PMCID: PMC8627561
- DOI: 10.1126/science.abg7071
Regulation of biomolecular condensates by interfacial protein clusters
Abstract
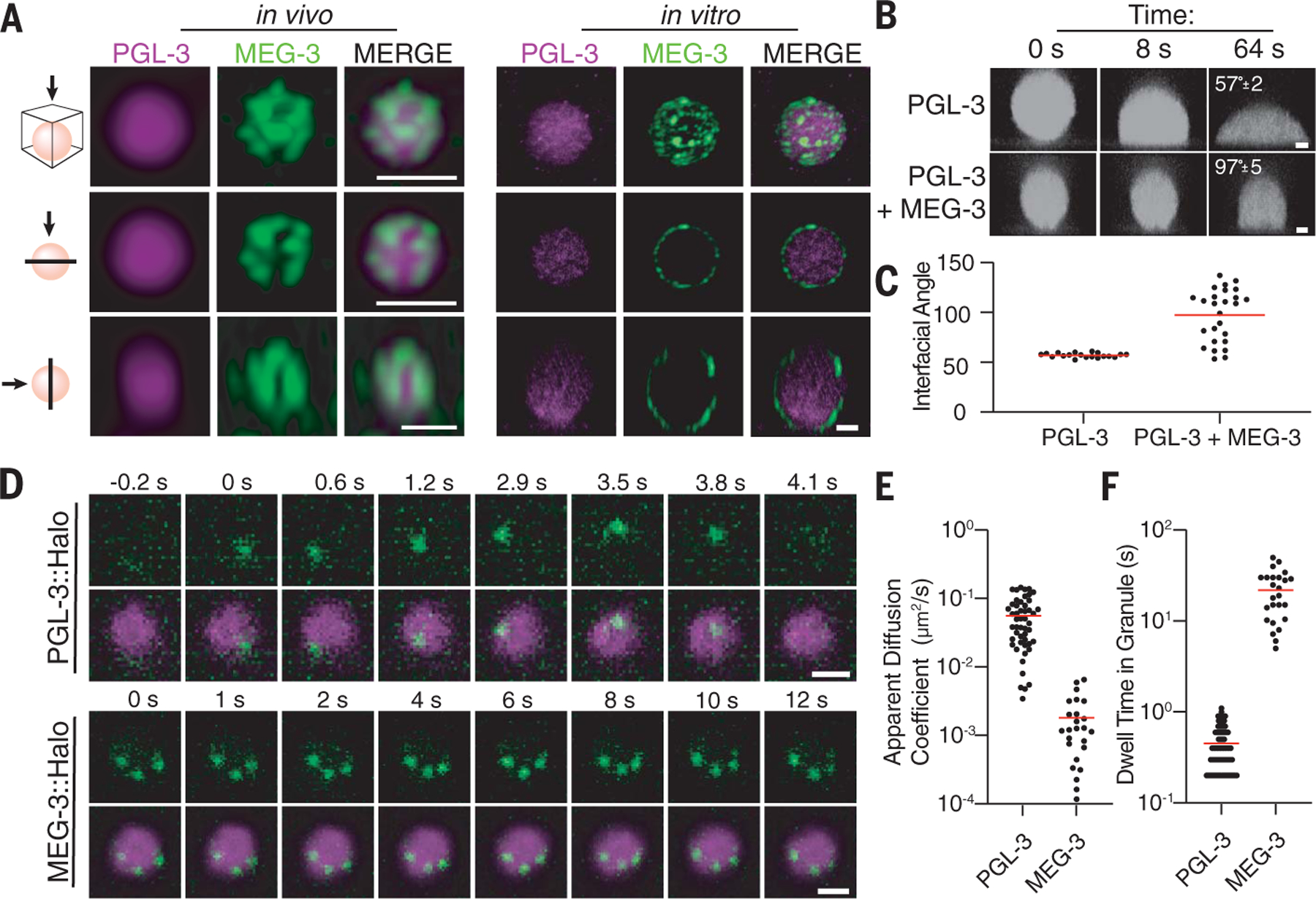
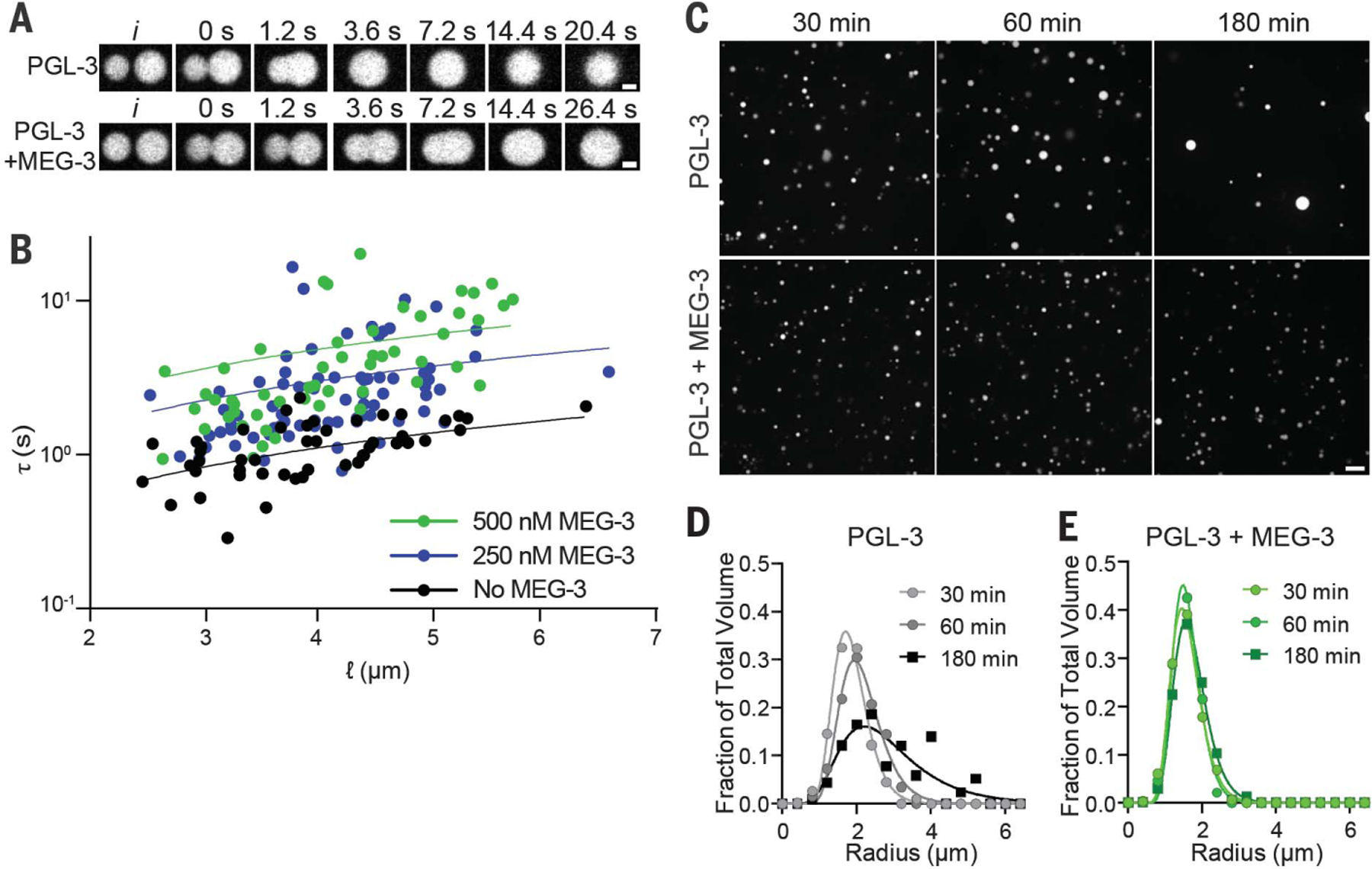
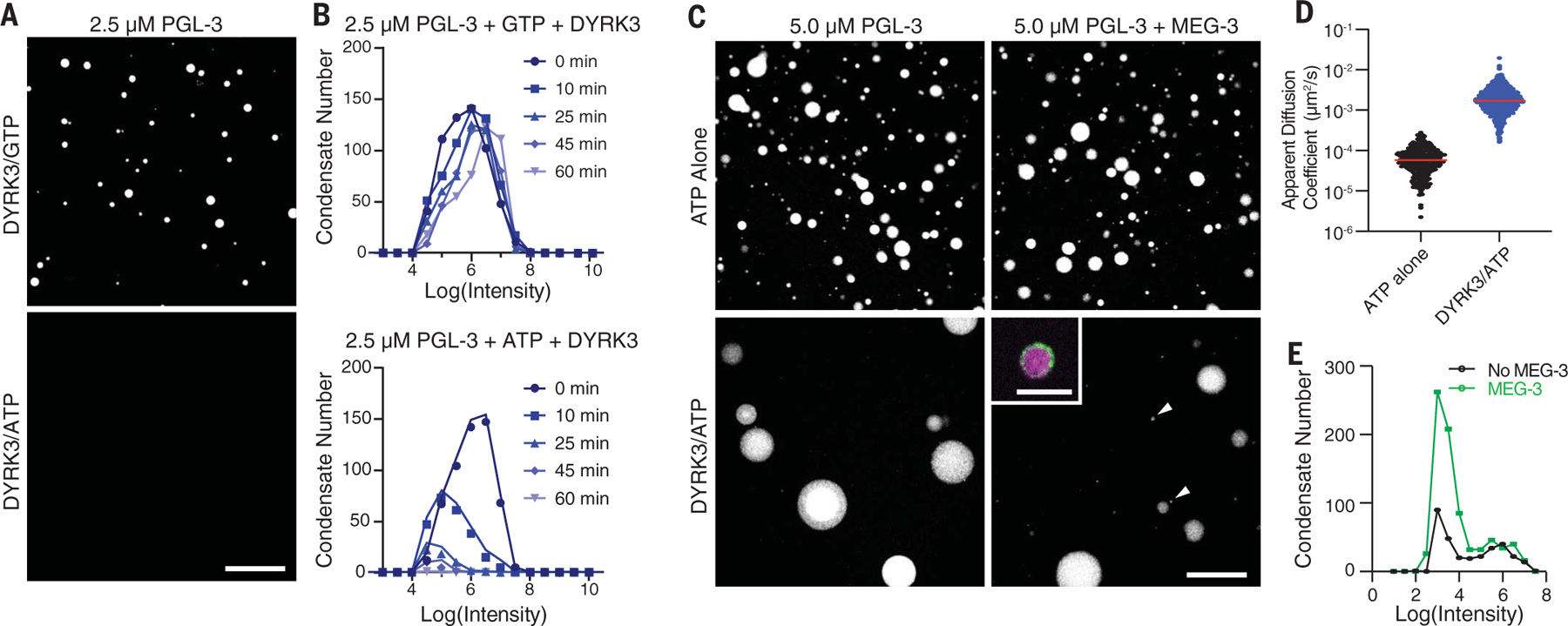
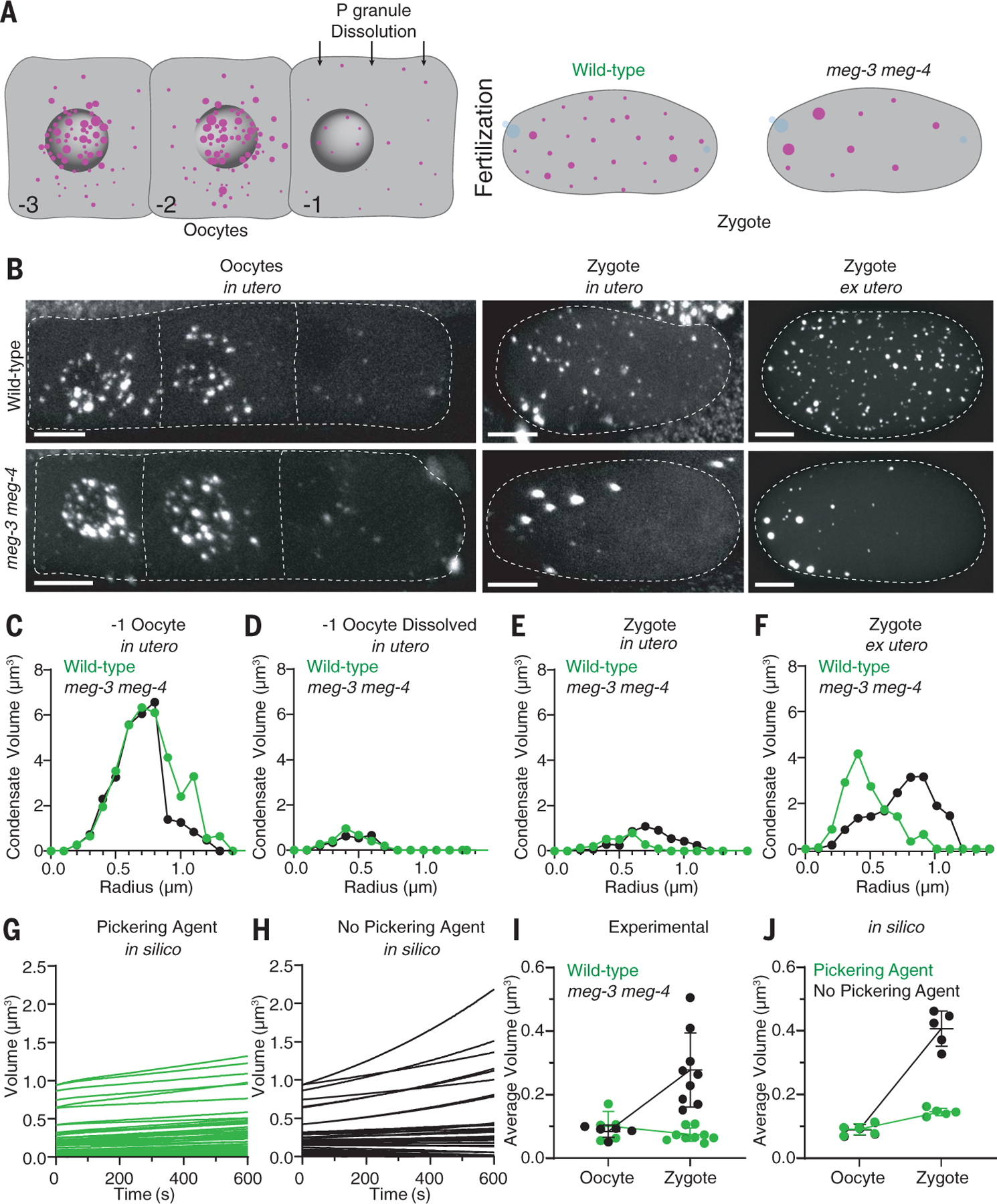
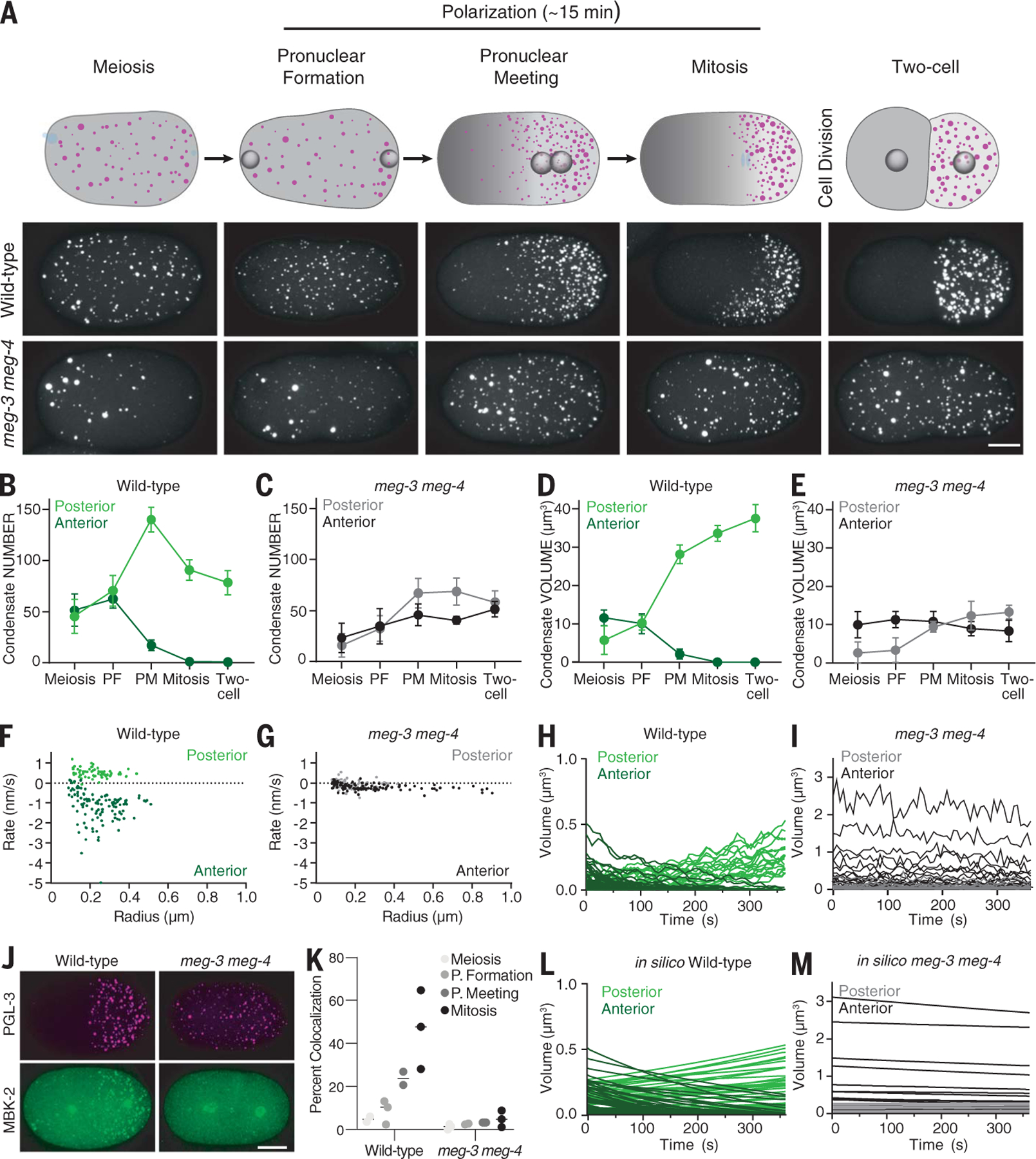
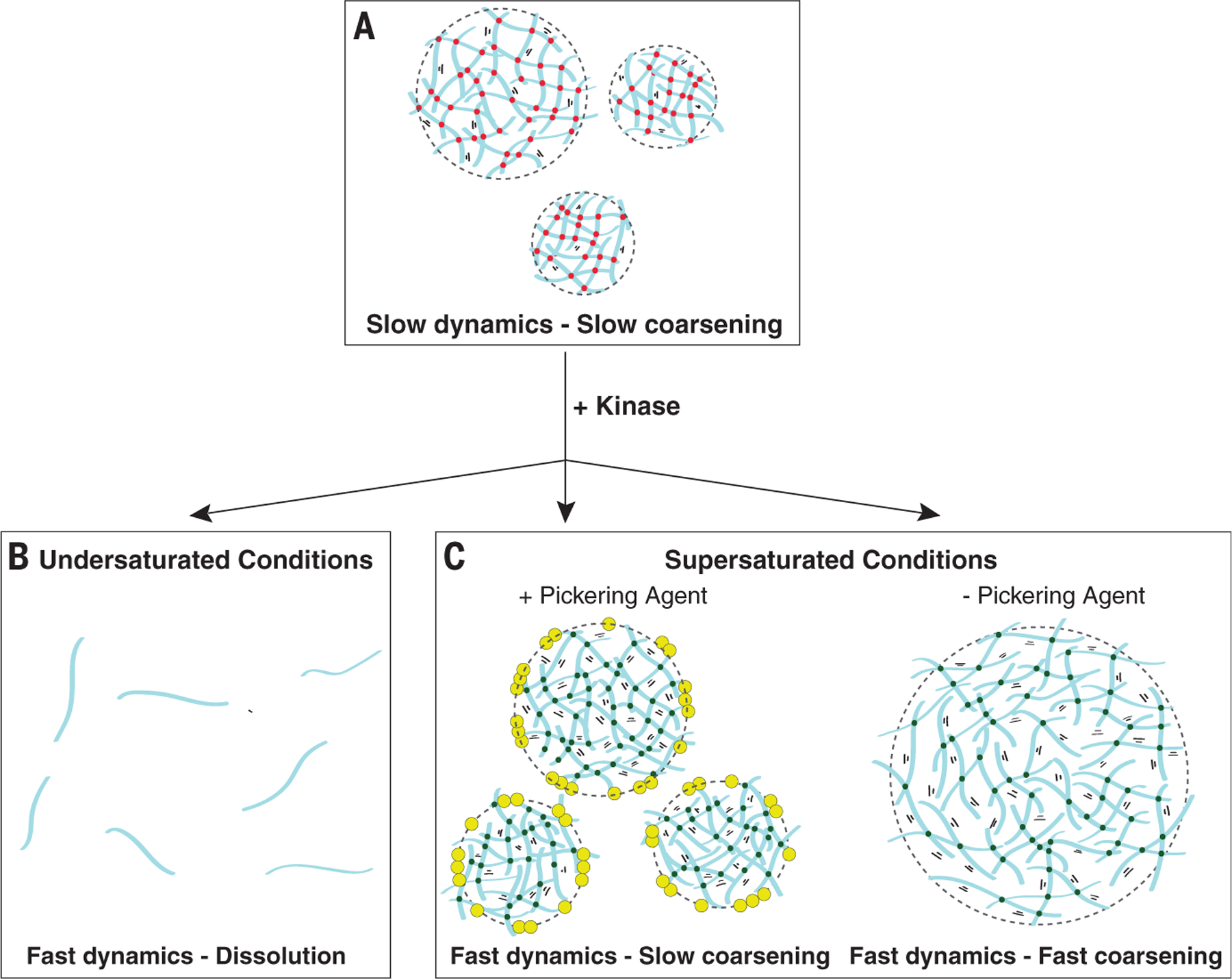
Biomolecular condensates are cellular compartments that can form by phase separation in the absence of limiting membranes. Studying the P granules of Caenorhabditis elegans, we find that condensate dynamics are regulated by protein clusters that adsorb to the condensate interface. Using in vitro reconstitution, live observations, and theory, we demonstrate that localized assembly of P granules is controlled by MEG-3, an intrinsically disordered protein that forms low dynamic assemblies on P granules. Following classic Pickering emulsion theory, MEG-3 clusters lower surface tension and slow down coarsening. During zygote polarization, MEG-3 recruits the DYRK family kinase MBK-2 to accelerate spatially regulated growth of the P granule emulsion. By tuning condensate-cytoplasm exchange, interfacial clusters regulate the structural integrity of biomolecular condensates, reminiscent of the role of lipid bilayers in membrane-bound organelles.
Conflict of interest statement
Figures
Comment in
-
Getting droplets into shape.Science. 2021 Sep 10;373(6560):1198-1199. doi: 10.1126/science.abl6282. Epub 2021 Sep 9. Science. 2021. PMID: 34516805
-
Signals from the interface: protein nanoclusters stabilize biomolecular condensates.Signal Transduct Target Ther. 2022 Jan 13;7(1):16. doi: 10.1038/s41392-022-00876-5. Signal Transduct Target Ther. 2022. PMID: 35027537 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
