

Architectures of neuronal circuits
- PMID: 34516844
- PMCID: PMC8916593
- DOI: 10.1126/science.abg7285
Architectures of neuronal circuits
Abstract
Although individual neurons are the basic unit of the nervous system, they process information by working together in neuronal circuits with specific patterns of synaptic connectivity. Here, I review common circuit motifs and architectural plans used in diverse brain regions and animal species. I also consider how these circuit architectures assemble during development and might have evolved. Understanding how specific patterns of synaptic connectivity can implement specific neural computations will help to bridge the huge gap between the biology of the individual neuron and the function of the entire brain, allow us to better understand the neural basis of behavior, and may inspire new advances in artificial intelligence.
Conflict of interest statement
Figures
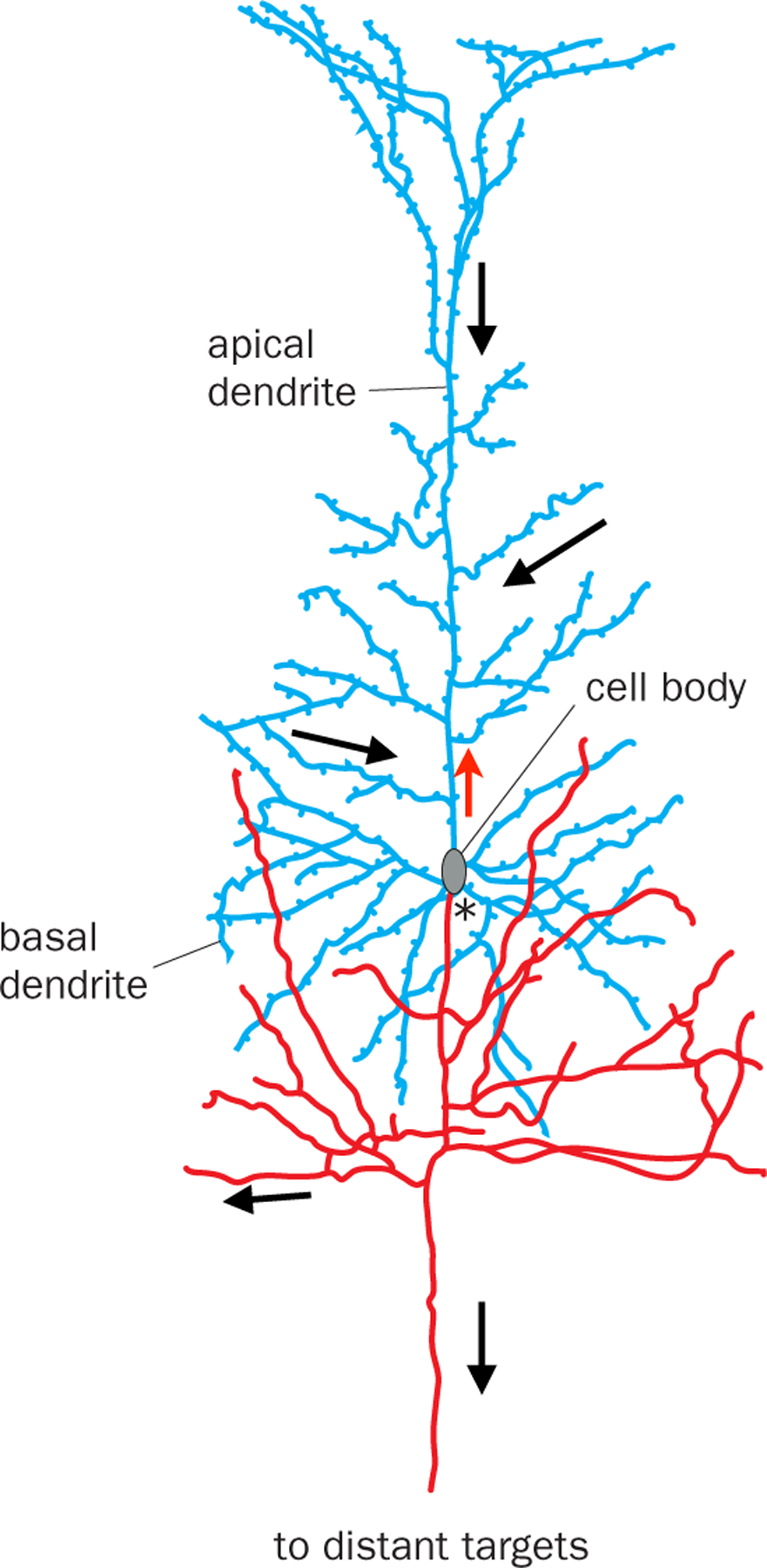
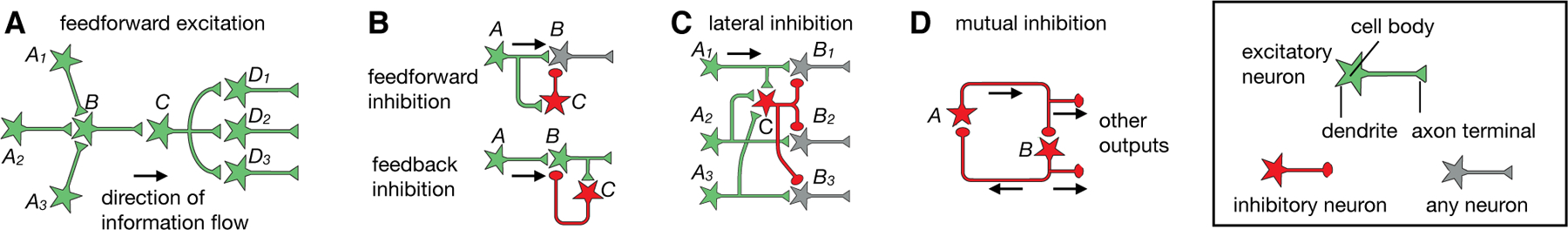
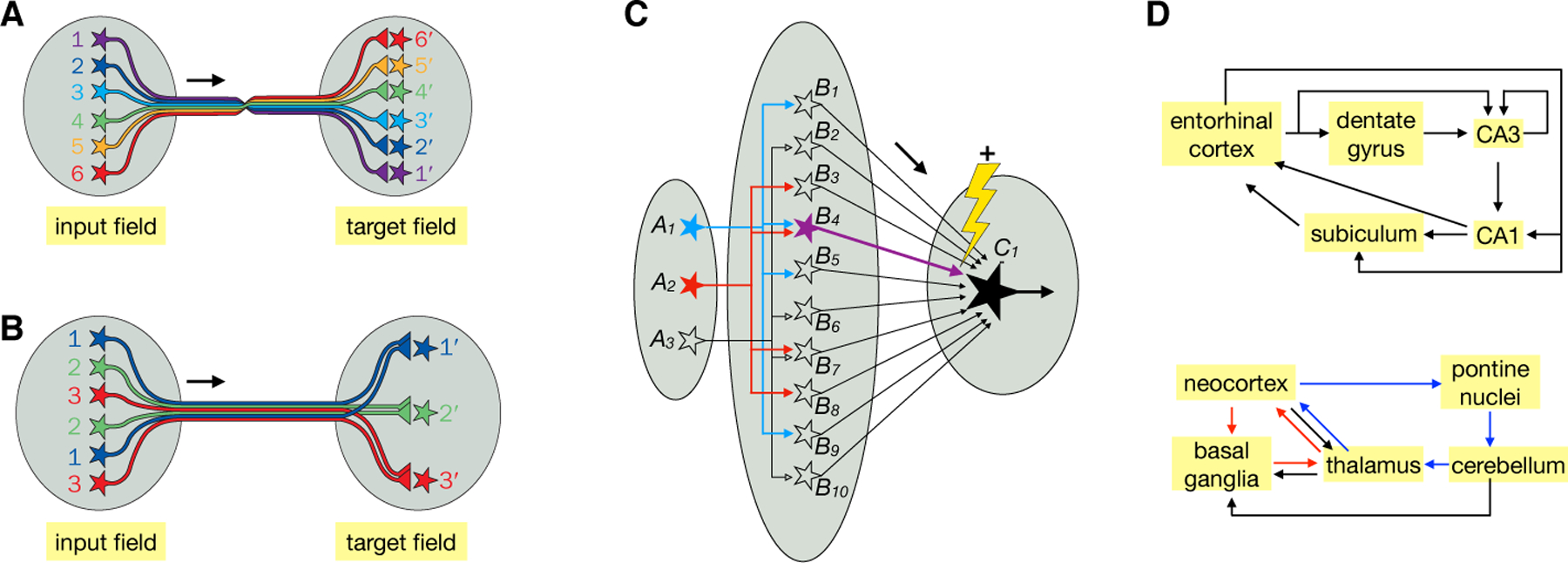
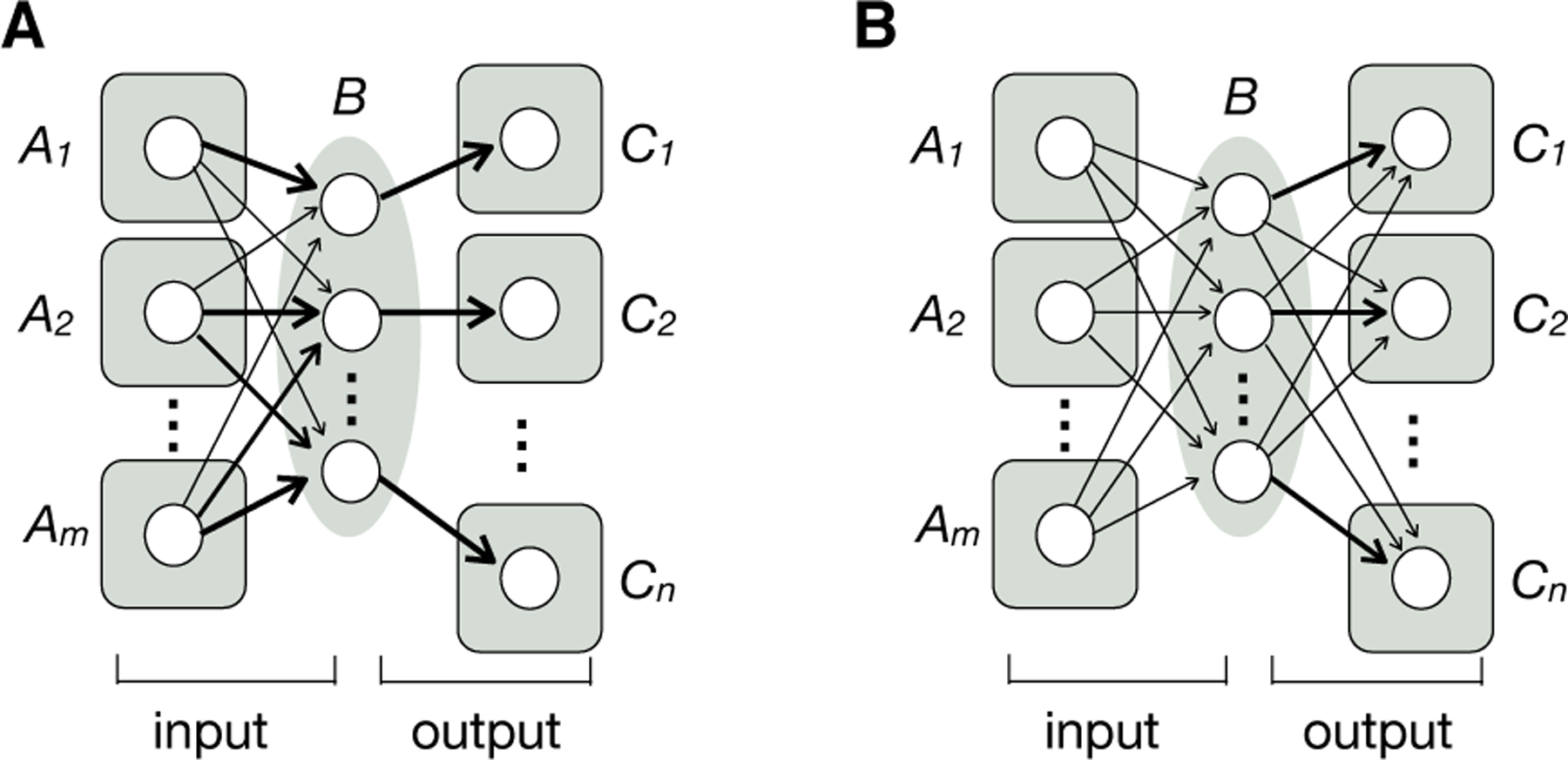
References
-
- Cajal SR, Histology of the Nervous System of Man and Vertebrates (Oxford University Press, Inc., translation by Swanson N & Swanson LW, Oxford, 1995).
-
- Golgi C, Sulla struttura della sostanza grigia del cervello. Gazetta medica lombarda IV, (1873).
-
- Laurent G et al. , Odor encoding as an active, dynamical process: Experiments, computation, and theory. Annu Rev Neurosci 24, 263–297 (2001). 11283312 - PubMed
-
- Shenoy KV, Sahani M, Churchland MM, Cortical control of arm movements: A dynamical systems perspective. Annu Rev Neurosci 36, 337–359 (2013). 23725001 - PubMed
-
- Luo L, Principles of Neurobiology, 2nd ed. (CRC Press/Garland Science, 2020).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
