

Regulation of protein function and degradation by heme, heme responsive motifs, and CO
- PMID: 34517731
- PMCID: PMC8966953
- DOI: 10.1080/10409238.2021.1961674
Regulation of protein function and degradation by heme, heme responsive motifs, and CO
Abstract
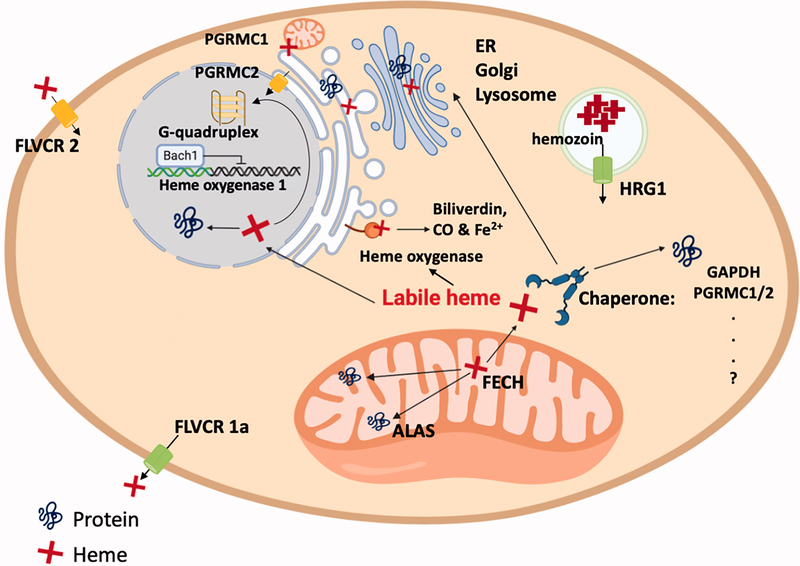
Heme is an essential biomolecule and cofactor involved in a myriad of biological processes. In this review, we focus on how heme binding to heme regulatory motifs (HRMs), catalytic sites, and gas signaling molecules as well as how changes in the heme redox state regulate protein structure, function, and degradation. We also relate these heme-dependent changes to the affected metabolic processes. We center our discussion on two HRM-containing proteins: human heme oxygenase-2, a protein that binds and degrades heme (releasing Fe2+ and CO) in its catalytic core and binds Fe3+-heme at HRMs located within an unstructured region of the enzyme, and the transcriptional regulator Rev-erbβ, a protein that binds Fe3+-heme at an HRM and is involved in CO sensing. We will discuss these and other proteins as they relate to cellular heme composition, homeostasis, and trafficking. In addition, we will discuss the HRM-containing family of proteins and how the stability and activity of these proteins are regulated in a dependent manner through the HRMs. Then, after reviewing CO-mediated protein regulation of heme proteins, we turn our attention to the involvement of heme, HRMs, and CO in circadian rhythms. In sum, we stress the importance of understanding the various roles of heme and the distribution of the different heme pools as they relate to the heme redox state, CO, and heme binding affinities.
Keywords: CO; Heme; circadian rhythm; enzymology; gas sensing/signaling; heme oxygenase; heme responsive motif; iron homeostasis; nuclear receptor; redox.
Conflict of interest statement
Disclosure statement
No potential conflict of interest was reported by the author(s).
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources