

Breast cancer-derived GM-CSF regulates arginase 1 in myeloid cells to promote an immunosuppressive microenvironment
- PMID: 34520398
- PMCID: PMC8516467
- DOI: 10.1172/JCI145296
Breast cancer-derived GM-CSF regulates arginase 1 in myeloid cells to promote an immunosuppressive microenvironment
Abstract
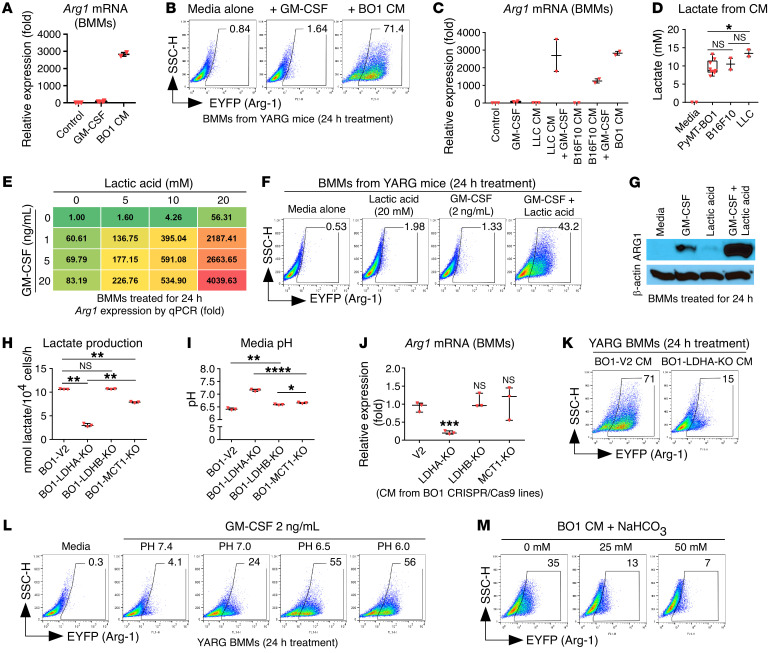
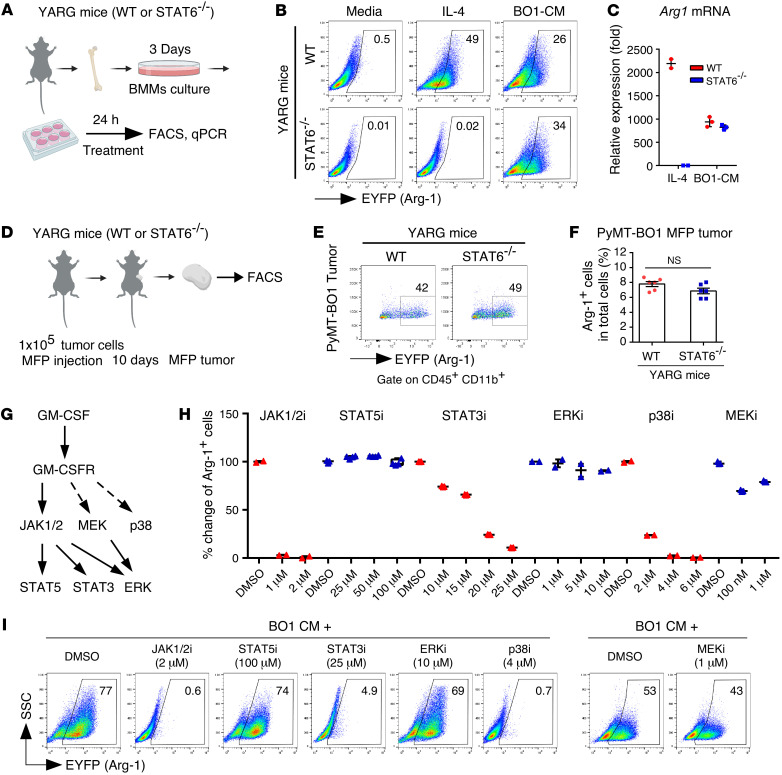
Tumor-infiltrating myeloid cells contribute to the development of the immunosuppressive tumor microenvironment. Myeloid cell expression of arginase 1 (ARG1) promotes a protumor phenotype by inhibiting T cell function and depleting extracellular l-arginine, but the mechanism underlying this expression, especially in breast cancer, is poorly understood. In breast cancer clinical samples and in our mouse models, we identified tumor-derived GM-CSF as the primary regulator of myeloid cell ARG1 expression and local immune suppression through a gene-KO screen of breast tumor cell-produced factors. The induction of myeloid cell ARG1 required GM-CSF and a low pH environment. GM-CSF signaling through STAT3 and p38 MAPK and acid signaling through cAMP were required to activate myeloid cell ARG1 expression in a STAT6-independent manner. Importantly, breast tumor cell-derived GM-CSF promoted tumor progression by inhibiting host antitumor immunity, driving a significant accumulation of ARG1-expressing myeloid cells compared with lung and melanoma tumors with minimal GM-CSF expression. Blockade of tumoral GM-CSF enhanced the efficacy of tumor-specific adoptive T cell therapy and immune checkpoint blockade. Taken together, we show that breast tumor cell-derived GM-CSF contributes to the development of the immunosuppressive breast cancer microenvironment by regulating myeloid cell ARG1 expression and can be targeted to enhance breast cancer immunotherapy.
Keywords: Breast cancer; Cancer immunotherapy; Immunology; Macrophages; Oncology.
Conflict of interest statement
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
- R21 AR073507/AR/NIAMS NIH HHS/United States
- T32 CA113275/CA/NCI NIH HHS/United States
- P30 AR074992/AR/NIAMS NIH HHS/United States
- S10 RR027552/RR/NCRR NIH HHS/United States
- R01 CA216840/CA/NCI NIH HHS/United States
- R01 AR070030/AR/NIAMS NIH HHS/United States
- P30 CA091842/CA/NCI NIH HHS/United States
- T32 CA009056/CA/NCI NIH HHS/United States
- R01 CA177670/CA/NCI NIH HHS/United States
- P30 AR057235/AR/NIAMS NIH HHS/United States
- P30 DK052574/DK/NIDDK NIH HHS/United States
- P01 CA100730/CA/NCI NIH HHS/United States
- T32 GM007200/GM/NIGMS NIH HHS/United States
- U54 CA199092/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
