

Tetraspanin 6 is a regulator of carcinogenesis in colorectal cancer
- PMID: 34521767
- PMCID: PMC8488650
- DOI: 10.1073/pnas.2011411118
Tetraspanin 6 is a regulator of carcinogenesis in colorectal cancer
Abstract
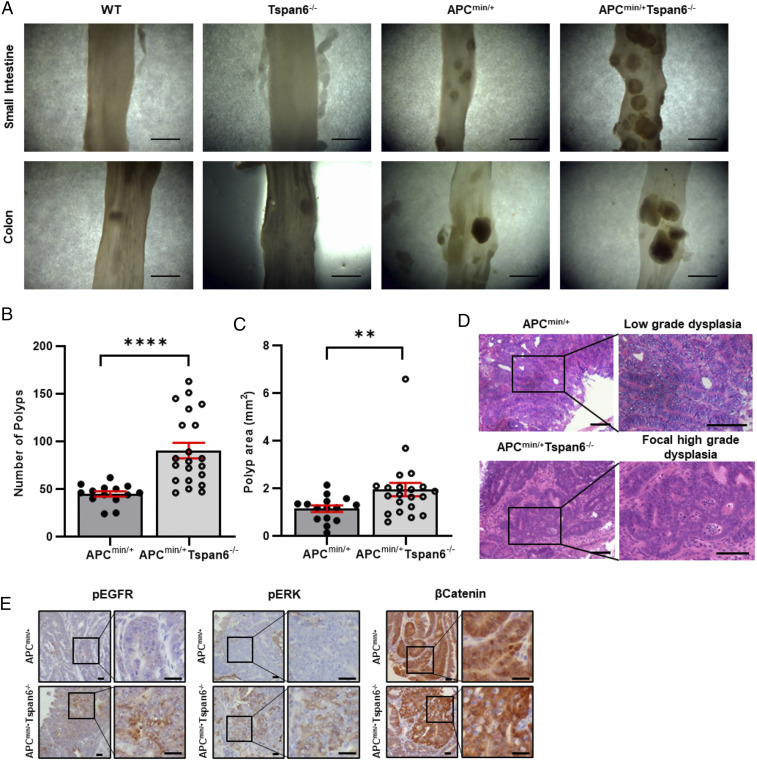
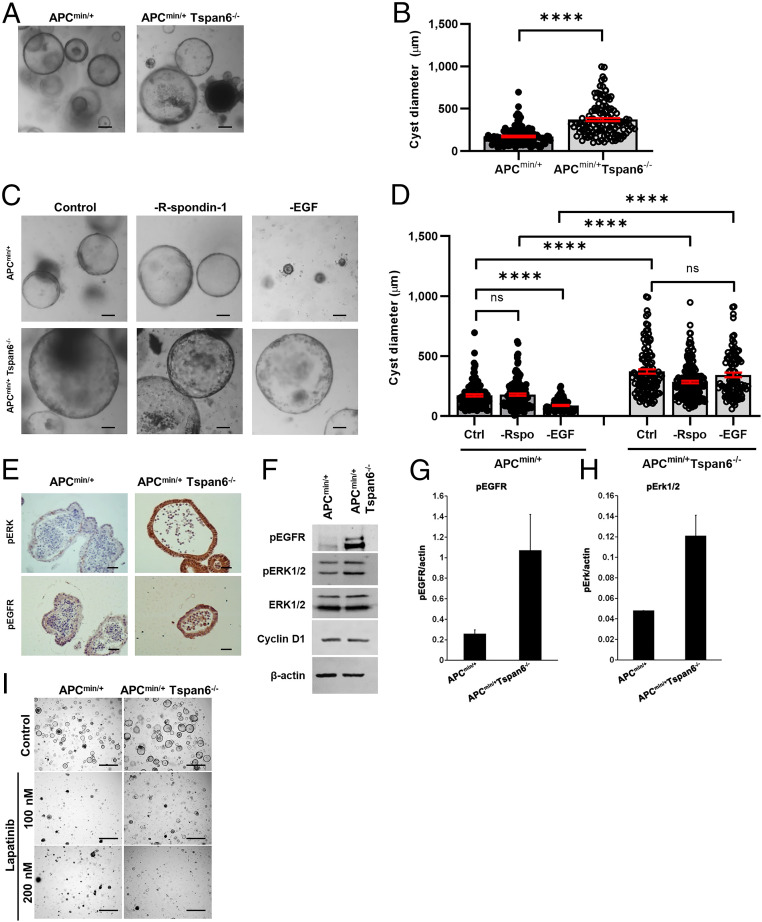
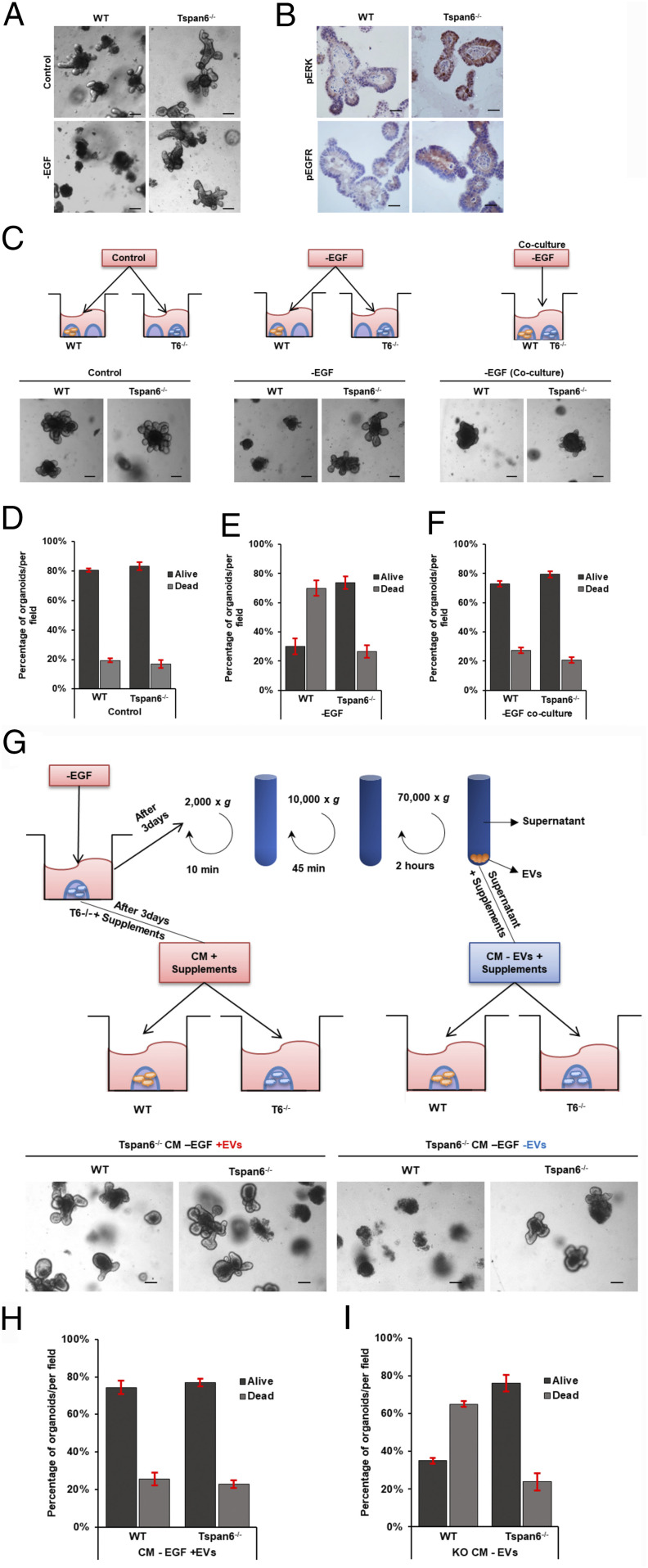
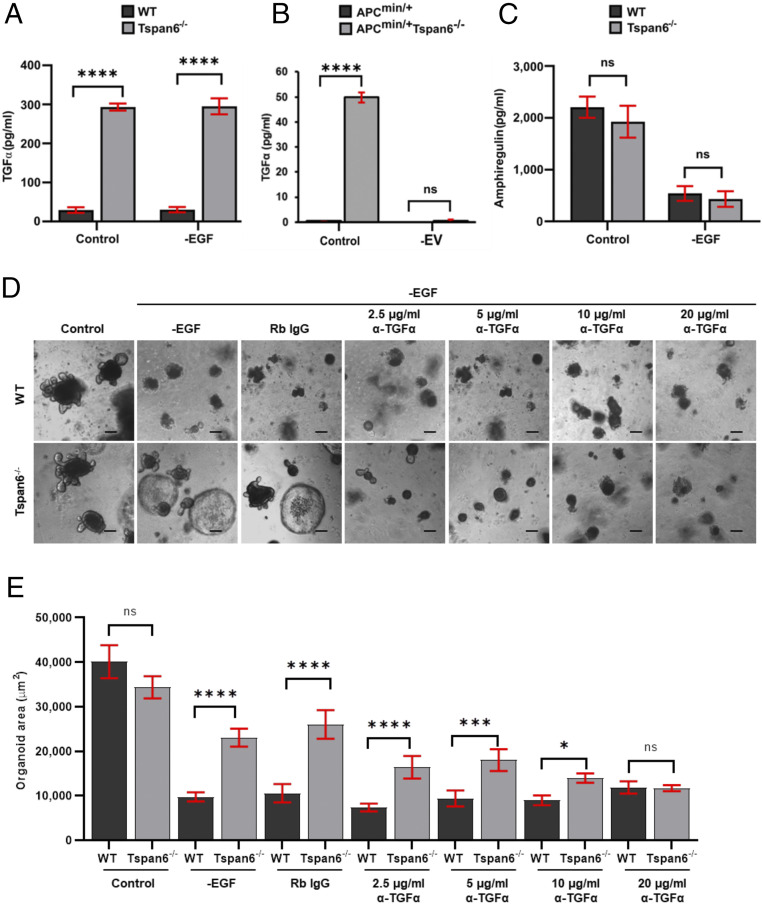
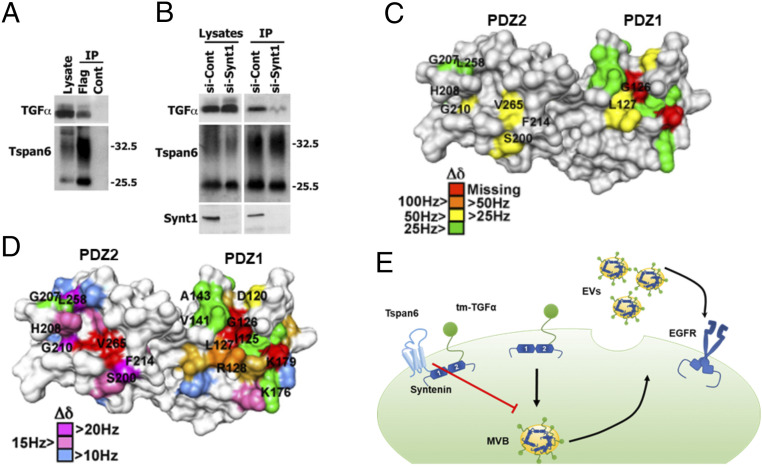
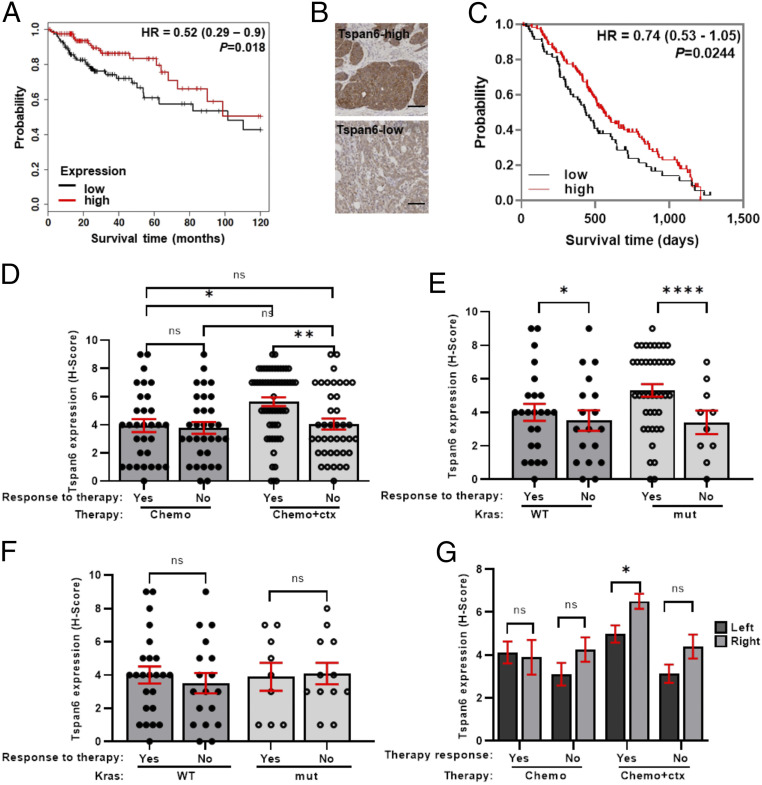
Early stages of colorectal cancer (CRC) development are characterized by a complex rewiring of transcriptional networks resulting in changes in the expression of multiple genes. Here, we demonstrate that the deletion of a poorly studied tetraspanin protein Tspan6 in Apcmin/+ mice, a well-established model for premalignant CRC, resulted in increased incidence of adenoma formation and tumor size. We demonstrate that the effect of Tspan6 deletion results in the activation of EGF-dependent signaling pathways through increased production of the transmembrane form of TGF-α (tmTGF-α) associated with extracellular vesicles. This pathway is modulated by an adaptor protein syntenin-1, which physically links Tspan6 and tmTGF-α. In support of this, the expression of Tspan6 is frequently decreased or lost in CRC, and this correlates with poor survival. Furthermore, the analysis of samples from the epidermal growth factor receptor (EGFR)-targeting clinical trial (COIN trial) has shown that the expression of Tspan6 in CRC correlated with better patient responses to EGFR-targeted therapy involving Cetuximab. Importantly, Tspan6-positive patients with tumors in the proximal colon (right-sided) and those with KRAS mutations had a better response to Cetuximab than the patients that expressed low Tspan6 levels. These results identify Tspan6 as a regulator of CRC development and a potential predictive marker for EGFR-targeted therapies in CRC beyond RAS pathway mutations.
Keywords: APCmin; EGFR; TGF alpha; colorectal cancer; tetraspanin.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Kaprio T., Hagström J., Andersson L. C., Haglund C., Tetraspanin CD63 independently predicts poor prognosis in colorectal cancer. Histol. Histopathol. 35, 887–892 (2020). - PubMed
-
- Greco C., et al. ., E-cadherin/p120-catenin and tetraspanin Co-029 cooperate for cell motility control in human colon carcinoma. Cancer Res. 70, 7674–7683 (2010). - PubMed
-
- Romanska H. M., Berditchevski F., Tetraspanins in human epithelial malignancies. J. Pathol. 223, 4–14 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
