

Dynamic reconfiguration of pro-apoptotic BAK on membranes
- PMID: 34523147
- PMCID: PMC8521275
- DOI: 10.15252/embj.2020107237
Dynamic reconfiguration of pro-apoptotic BAK on membranes
Abstract
BAK and BAX, the effectors of intrinsic apoptosis, each undergo major reconfiguration to an activated conformer that self-associates to damage mitochondria and cause cell death. However, the dynamic structural mechanisms of this reconfiguration in the presence of a membrane have yet to be fully elucidated. To explore the metamorphosis of membrane-bound BAK, we employed hydrogen-deuterium exchange mass spectrometry (HDX-MS). The HDX-MS profile of BAK on liposomes comprising mitochondrial lipids was consistent with known solution structures of inactive BAK. Following activation, HDX-MS resolved major reconfigurations in BAK. Mutagenesis guided by our HDX-MS profiling revealed that the BCL-2 homology (BH) 4 domain maintains the inactive conformation of BAK, and disrupting this domain is sufficient for constitutive BAK activation. Moreover, the entire N-terminal region preceding the BAK oligomerisation domains became disordered post-activation and remained disordered in the activated oligomer. Removal of the disordered N-terminus did not impair, but rather slightly potentiated, BAK-mediated membrane permeabilisation of liposomes and mitochondria. Together, our HDX-MS analyses reveal new insights into the dynamic nature of BAK activation on a membrane, which may provide new opportunities for therapeutic targeting.
Keywords: BAK; BCL-2; apoptosis; hydrogen-deuterium exchange mass spectrometry; membrane.
© 2021 The Authors.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
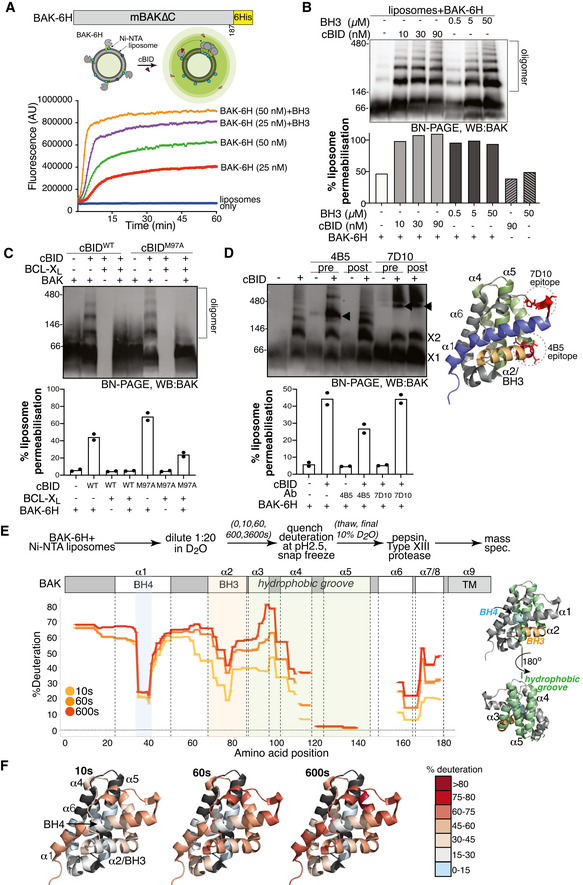
BAK‐6H permeabilising activity is exacerbated by cBID. BAK‐6H (mBAK∆C21‐6H) was loaded at the indicated concentrations onto Ni‐NTA liposomes (5 µM) prior to the addition of cBID. The release of carboxyfluorescein was measured over time. Results representative of at least three independent experiments.
BAK‐6H oligomerises on liposomes induced by cBID. Liposomes were incubated with BAK‐6H (150 nM) and the indicated concentration of cBID or a BID BH3 peptide for 60 min. Liposomes were solubilised in digitonin and BAK oligomers were analysed by BN‐PAGE. In parallel, samples were tested at endpoint for liposome permeabilisation based on the release of fluorescent dextran, normalised to total fluorescence of detergent permeabilised liposomes. Results representative of three independent experiments.
BAK‐6H oligomerisation on liposomes is blocked by BCL‐XL. Liposomes were incubated with BAK‐6H (150 nM) together with cBID (WT, 90 nM) or cBID M97A (90 nM) that has reduced affinity for BCL‐XL, in the presence or absence of BCL‐XL for 60 min prior to analysis of BAK oligomerisation by BN‐PAGE. Results representative of two independent experiments. Liposome permeabilisation was monitored with the indicated combinations of cBID (90 nM), BAK‐6H (150 nM) and BCL‐XL (1 µM). Data are presented as mean of two independent experiments with percentage release relative to detergent‐lysed liposomes at a timepoint when permeabilisation with BAK and cBid was approximately 50% of detergent‐lysis maximum.
BAK‐6H oligomerisation on liposomes involves its BH3 domain. Liposomes were incubated with BAK‐6H (150 nM) together with cBID (90 nM). Antibody recognising the BAK BH3 domain (4B5) or α1‐α2 loop (7D10), as shown on the structure of BAK (PDB:2IMS), were added (2.4 µg/µl) either prior to (pre) or after (post) incubation with cBID for 60 min. BAK:Ab complex is indicated (arrowhead). 7D10 which recognises the activated form of mBAK (Iyer et al, 2016) bound to and gel‐shifted BAK when added either before or after activation with cBID. In contrast, 4B5 failed to appreciably gel‐shift inactive BAK (lanes 3 and 5) or BAK that was already activated (lane 6), but gel‐shifted BAK when present during cBID activation (lane 4). Liposome permeabilisation was assessed after 20 min incubation with cBID (90 nM), BAK‐6H (150 nM) and antibodies 4B5 (2.5 µg) or 7D10 (2.5 µg). Data are presented as mean of two independent experiments with percentage release relative to detergent‐lysed liposomes at a timepoint when permeabilisation with BAK and cBid was approximately 50% of detergent‐lysis maximum.
HDX‐MS reveals structural constraints of BAK on a membrane. BAK‐6H (150 nM) on liposomes was deuterated for 0, 10s, 60s or 600s and the incorporation of deuterium was assessed by mass spectrometry. Deuteration is expressed relative to a theoretical maximum. BH3 domain (orange), BH4 domain (blue) and hydrophobic surface groove comprising αhelices 3–5 (green) are shown on the structure of BAK (PDB:2IMS).
Differential deuteration of BAK in a membrane. Relative deuteration is mapped onto the structure of inactive monomeric BAK (2IMS, Moldoveanu et al, 2006).
- A, B
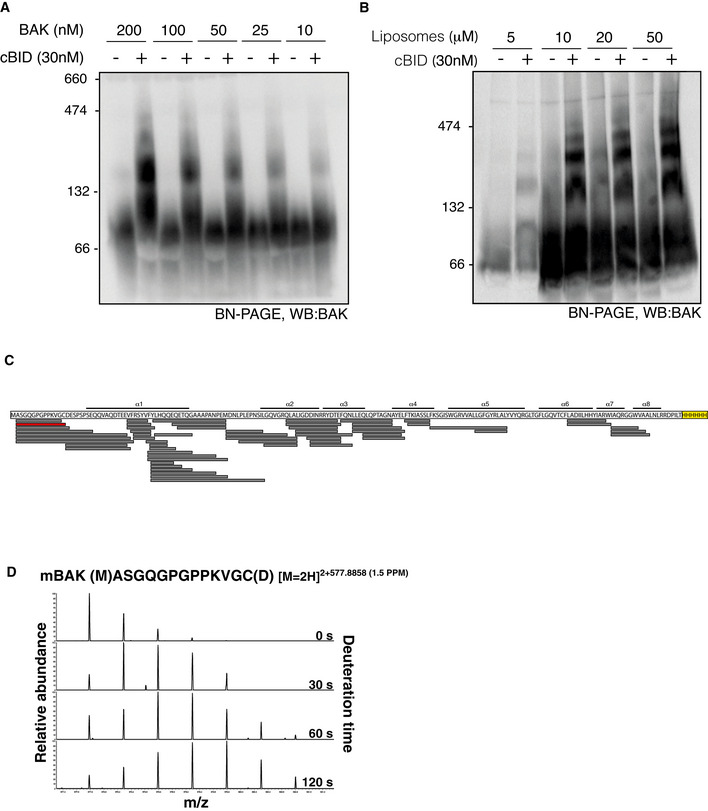
Liposomes were incubated with BAK‐6H and cBID at the indicated concentration prior to the analysis of BAK oligomerisation on BN‐PAGE.
- C
BAK peptide coverage following combined proteolysis with pepsin and Aspergillus Type XIII protease.
- D
Example of time‐dependent deuteration of BAK peptide. The increase in m/z of a BAK peptide (shown in red in C) due to incorporation with deuterium over time.
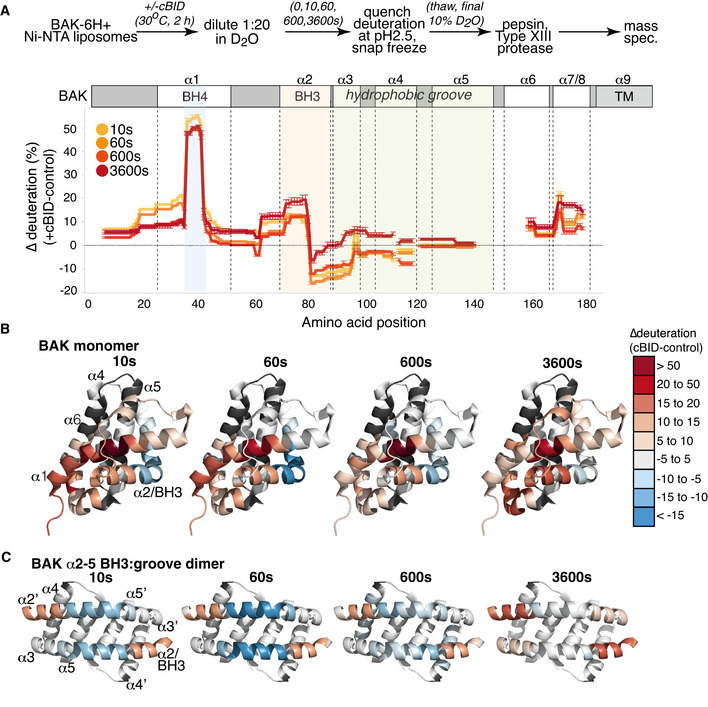
BAK‐6H on liposomes was activated or not with cBID for 60 min prior to deuteration for 10s, 60s, 600s, or 3,600s. Incorporation of deuterium was assessed by mass spectrometry. Change in deuteration at each time point is expressed relative to BAK on liposomes without cBID.
Deuteration of BAK in a membrane compared to inactive BAK is mapped onto the structure of inactive monomeric BAK (2IMS, Moldoveanu et al, 2006).
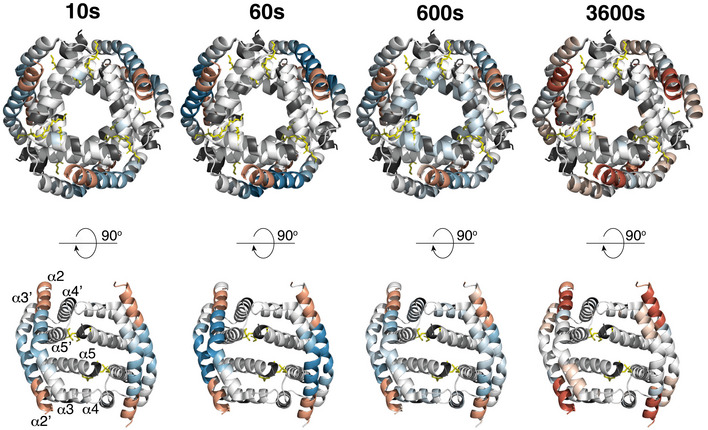
Deuteration of BAK in a membrane compared to inactive BAK is mapped onto the structure of an activated BAK BH3:groove dimer (4U2V, Brouwer et al, 2014).
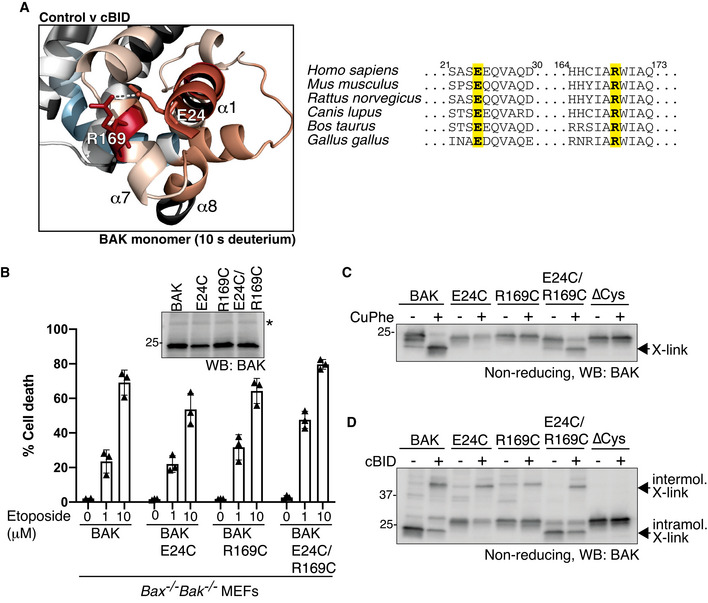
HDX‐MS reveals a significant increase in deuteration at positions E24 (α1) and R169 (α7) following activation with cBID. These positions are conserved suggesting a potentially important hydrogen bonding interaction.
BAK, or BAK with cysteine introduced at the indicated positions (on a BAK∆Cys background) were stably expressed in Bax −/− Bak −/− MEFs and assessed for BAK expression by immunoblotting (inset panel; * indicates non‐specific band) and apoptotic activity in response to etoposide treatment. Data are mean ± SD of three independent experiments.
E24 and R169 are proximal in inactive BAK on mitochondria. Mitochondria‐enriched membrane fractions from cells in (B) were incubated with oxidant (CuPhe) and induced intramolecular disulphide linkage of BAK was assessed on non‐reducing SDS–PAGE.
E24 and R169 dissociate during BAK activation. Mitochondria‐enriched membrane fractions from cells in (B) were incubated with cBID (100 nM) prior to oxidant (CuPhe) and the induced intramolecular and intermolecular disulphide linkage of BAK was assessed on non‐reducing SDS–PAGE.
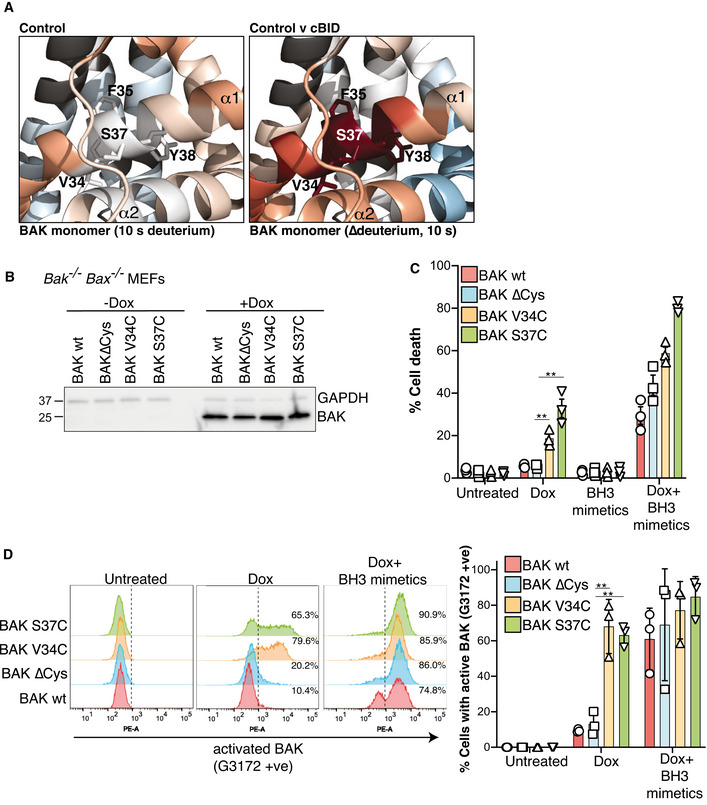
HDX‐MS reveals a constrained BH4 domain in inactive BAK that is rapidly deuterated after activation. BAK BH4 residues in the α1 (V34, F35, S37 and Y38) corresponding to the BH4 motif Φ1Φ2XXΦ3Φ4 (Kvansakul et al, 2008) are indicated as sticks.
BAK wild‐type or BAK engineered with the indicated mutations (on a BAK∆Cys background) were stably expressed in Bax −/− Bak −/− MEFs and expression was assessed by immunoblotting following 3 h doxycycline treatment.
Bax −/− Bak −/− MEFs expressing the indicated BAK variants were treated with doxycycline (Dox, 3 h), incubated for a further 24 h in the presence or absence of BH3 mimetic compounds, and cell death was assessed by PI uptake. Data are expressed as mean ± SD of three independent experiments, **P < 0.01, unpaired Student’s t‐test.
Mutation of BH4 promotes BAK activating conformation change. Cells in (B) were treated with doxycycline (3 h) to induce BAK expression followed by incubation with BH3 mimetics where indicated (for 2 h) and conformation change of BAK was assessed by intracellular flow cytometry with an antibody that recognises activated BAK (G3172). FACS profiles from one experiment are representative of the three independent experiments shown in Fig EV3. Collated data from three independent experiments are expressed as per cent G3172 positive cells (mean ± SD), **P < 0.01 unpaired Student’s t‐test.
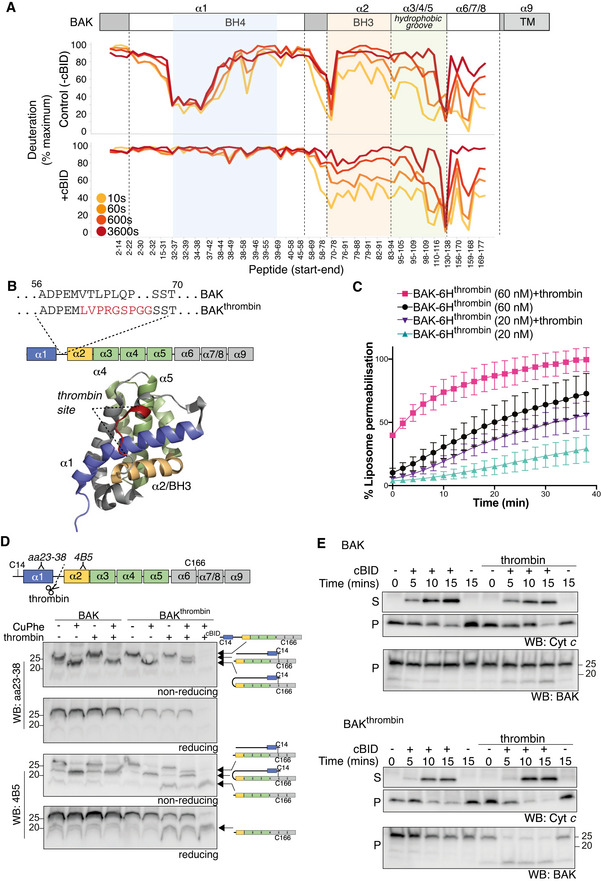
BAK‐6H on liposomes was activated (+cBID, bottom) or not (Control, top) with cBID for 60 min prior to deuteration for 10 s, 60 s, 600 s, or 3,600 s. Incorporation of deuterium was assessed by mass spectrometry and is expressed relative to a non‐deuterated sample and a theoretical maximum. HDX‐MS reveals that following activation with cBID, maximal deuteration of the BAK N‐terminus (up to amino acid 58) occurred within 10 secs.
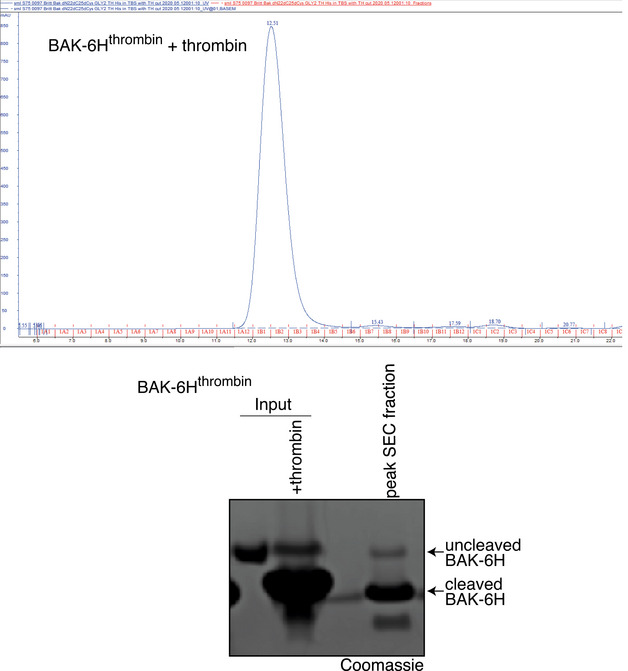
BAK mutant with an engineered thrombin cleavage site in α1‐α2 loop of hBAK∆N22/∆C25/C166S with a 6xHis C‐terminal tag for recombinant protein expression or as full‐length protein for ectopic expression in MEFs (BAKthrombin).
BAK cleaved within its α1‐2 loop by thrombin is able to mediate liposome permeabilisation. Recombinant BAK‐6Hthrombin pre‐cleaved or not with thrombin (see Fig EV4) was incubated with liposomes at either 20 or 60 nM and liposome permeabilisation induced by cBID was monitored over time. Data are presented as percentage liposome permeabilisation relative to detergent‐lysed liposomes, mean ± SEM of three independent experiments.
BAK wild‐type or BAK with an engineered α1‐α2 loop thrombin cleavage site were stably expressed in Bax −/− Bak −/− MEFs. Membrane fractions were incubated with thrombin prior to incubation with copper phenanthroline (CuPhe) to induce disulphide linkage. Samples were run on SDS–PAGE under non‐reducing or reducing conditions and immunoblotted for BAK with an antibody recognising the BAK N‐terminus upstream of the thrombin cleavage site (aa23–38) or an antibody recognising the BH3 domain (4B5) downstream of the cleavage site. Schematic indicates the nature of the BAK protein detected by each antibody. Disulphide linkage indicated by a hashed line. Note that the N‐terminal portion of thrombin cleaved BAK could not be detected with the N‐terminal antibody unless it was intramolecularly disulphide‐linked. Where indicated cBID (100 nM) was added to activate BAK during thrombin cleavage. Data representative of two independent experiments.
Cleavage of the BAK α1‐α2 loop potentiates MOMP. Membrane fractions from Bax −/− Bak −/− MEFs expressing BAK or BAKthrombin were incubated or not with thrombin in the presence or absence of cBID (10 nM) for the indicated times. Membrane (P) and supernatant (S) fractions were separated and immunoblotted for cytochrome c or BAK 4B5. Data representative of three independent experiments.
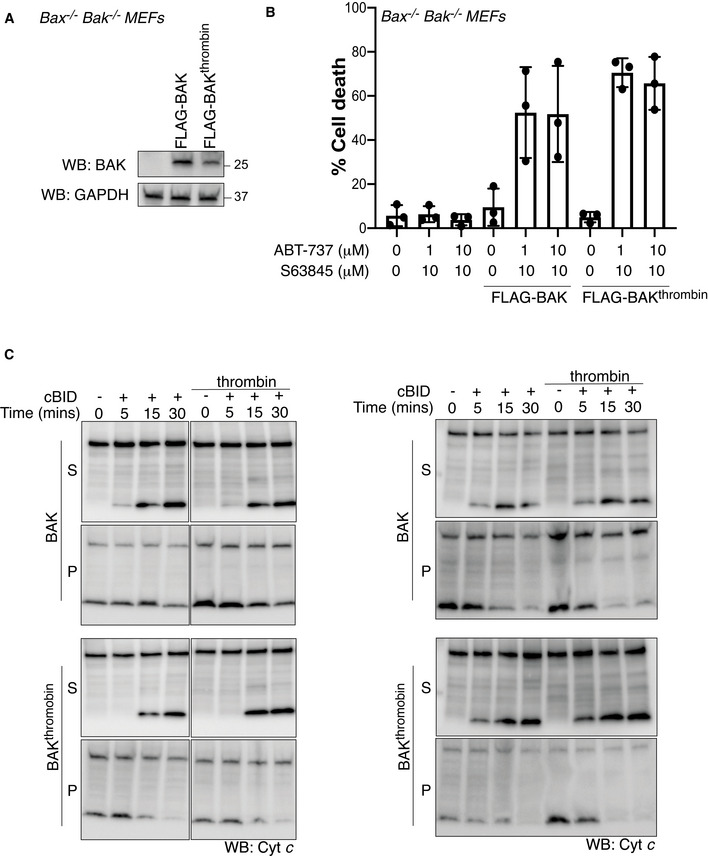
FLAG‐BAK or FLAG‐BAKthrombin were stably expressed in Bax −/− Bak −/− MEFs and assessed by immunoblotting.
Cells from A were treated with combined BH3 mimetic drugs for 24 h and cell death was analysed by PI uptake. Data are mean ± SD of three independent experiments.
Mitochondria‐enriched fractions isolated from cells in A were incubated with cBID (10 nM) in the presence or absence of thrombin for the indicated times and membrane pellet (P) and supernatant (S) were separated and immunoblotted for cytochrome c or BAK (4B5). Data from two independent experiments are shown, third experiment is shown in Fig 5E.
Comment in
-
A killer metamorphosis: catching BAK in action at the membrane.EMBO J. 2021 Oct 18;40(20):e109529. doi: 10.15252/embj.2021109529. Epub 2021 Sep 20. EMBO J. 2021. PMID: 34542920 Free PMC article.
References
-
- Alsop AE, Fennell SC, Bartolo RC, Tan IK, Dewson G, Kluck RM (2015) Dissociation of Bak alpha1 helix from the core and latch domains is required for apoptosis. Nat Commun 6: 6841 - PubMed
-
- Basanez G, Sharpe JC, Galanis J, Brandt TB, Hardwick JM, Zimmerberg J (2002) Bax‐type apoptotic proteins porate pure lipid bilayers through a mechanism sensitive to intrinsic monolayer curvature. J Biol Chem 277: 49360–49365 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
