

DNA Repair in Staphylococcus aureus
- PMID: 34523959
- PMCID: PMC8483670
- DOI: 10.1128/MMBR.00091-21
DNA Repair in Staphylococcus aureus
Abstract
Staphylococcus aureus is a common cause of both superficial and invasive infections of humans and animals. Despite a potent host response and apparently appropriate antibiotic therapy, staphylococcal infections frequently become chronic or recurrent, demonstrating a remarkable ability of S. aureus to withstand the hostile host environment. There is growing evidence that staphylococcal DNA repair makes important contributions to the survival of the pathogen in host tissues, as well as promoting the emergence of mutants that resist host defenses and antibiotics. While much of what we know about DNA repair in S. aureus is inferred from studies with model organisms, the roles of specific repair mechanisms in infection are becoming clear and differences with Bacillus subtilis and Escherichia coli have been identified. Furthermore, there is growing interest in staphylococcal DNA repair as a target for novel therapeutics that sensitize the pathogen to host defenses and antibiotics. In this review, we discuss what is known about staphylococcal DNA repair and its role in infection, examine how repair in S. aureus is similar to, or differs from, repair in well-characterized model organisms, and assess the potential of staphylococcal DNA repair as a novel therapeutic target.
Keywords: DNA damage; DNA repair; SOS; Staphylococcus aureus; antibiotic resistance; bacteriophage; neutrophil.
Figures

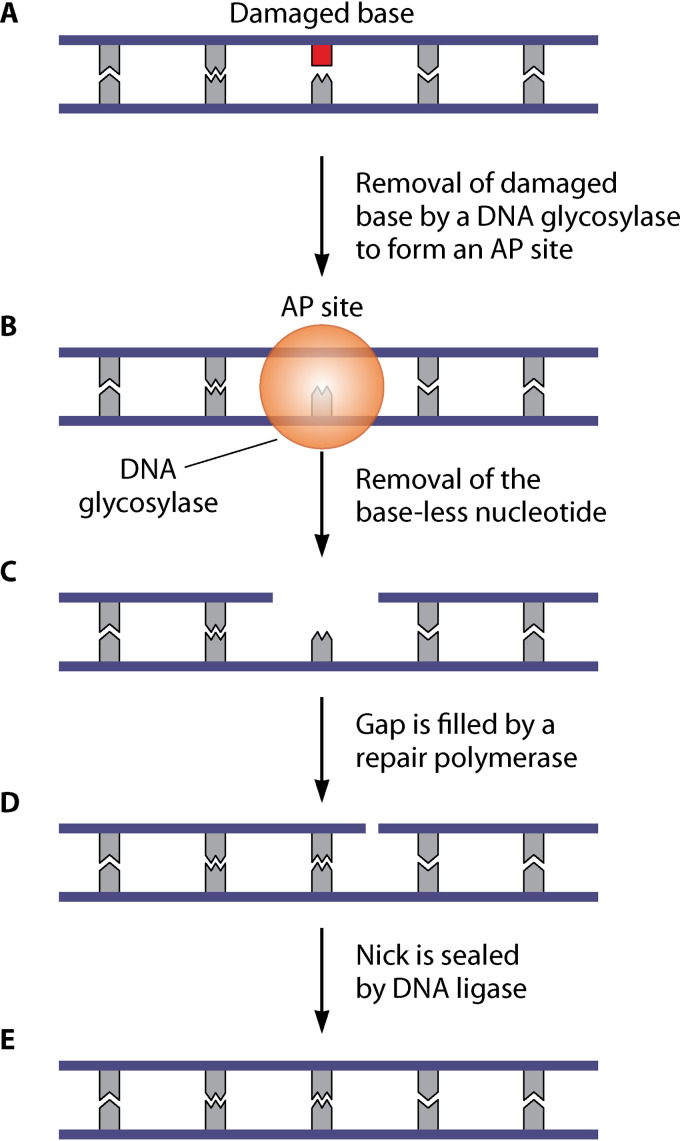


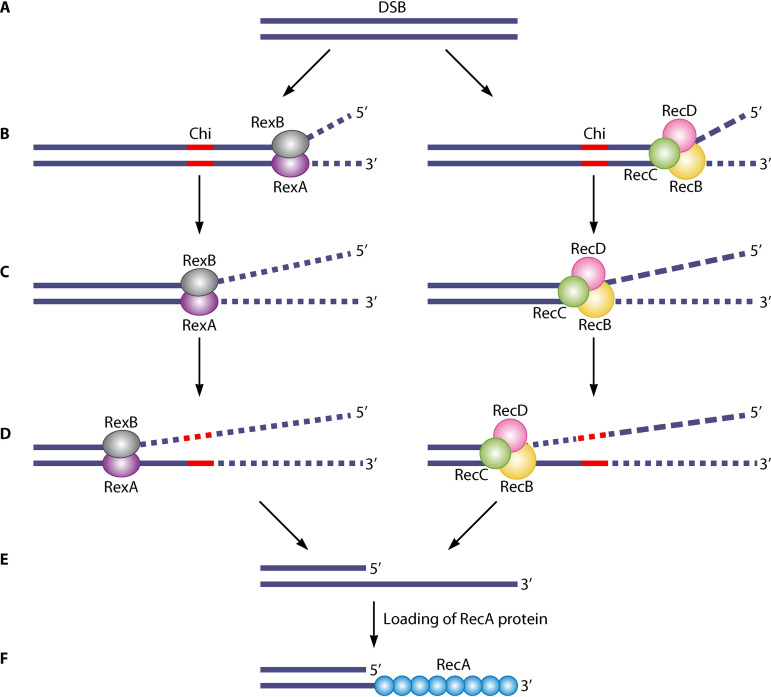

References
-
- Friedberg EC, Wood RD, Walker GC, Schultz RA, Siede W, Ellenberger T. 2006. DNA repair and mutagenesis. American Society for Microbiology Press, Washington, DC.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
