

Olfactory Entry Promotes Herpesvirus Recombination
- PMID: 34523965
- PMCID: PMC8577388
- DOI: 10.1128/JVI.01555-21
Olfactory Entry Promotes Herpesvirus Recombination
Abstract
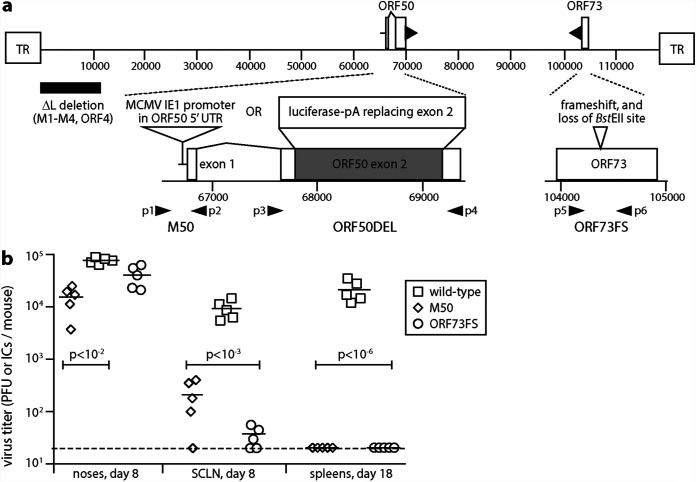
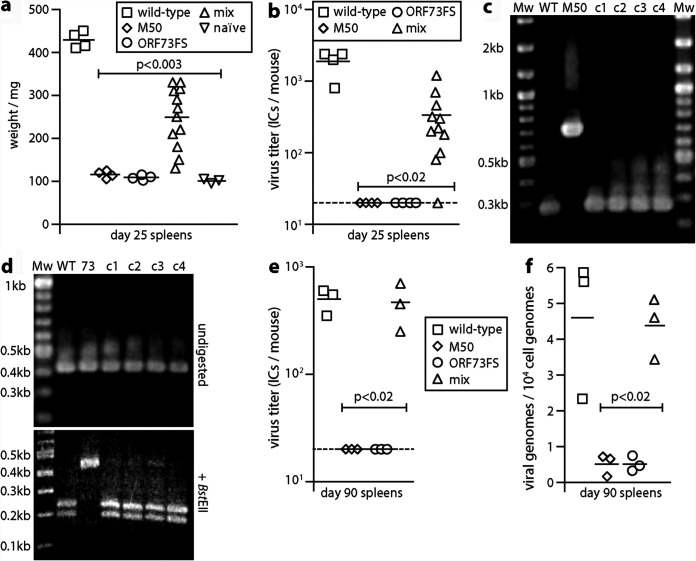
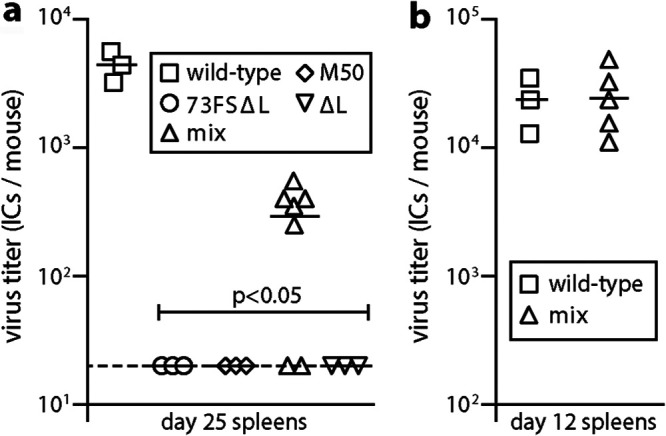
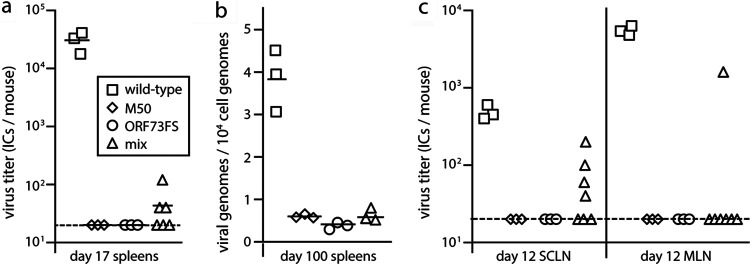
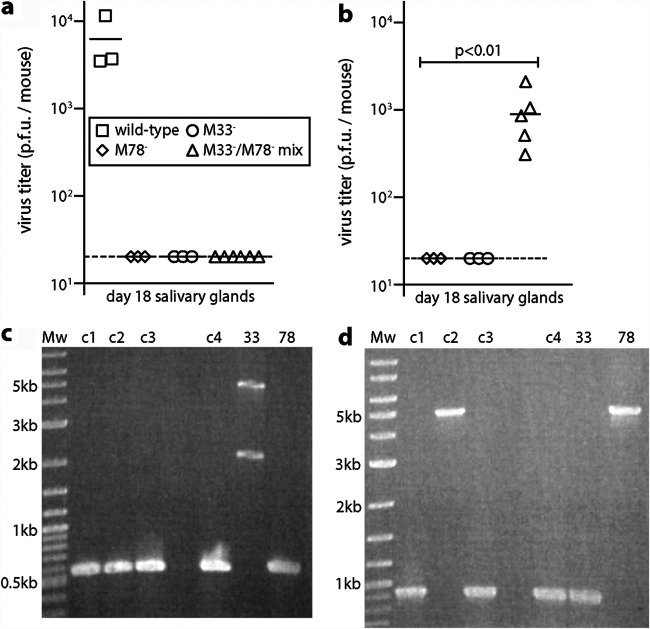
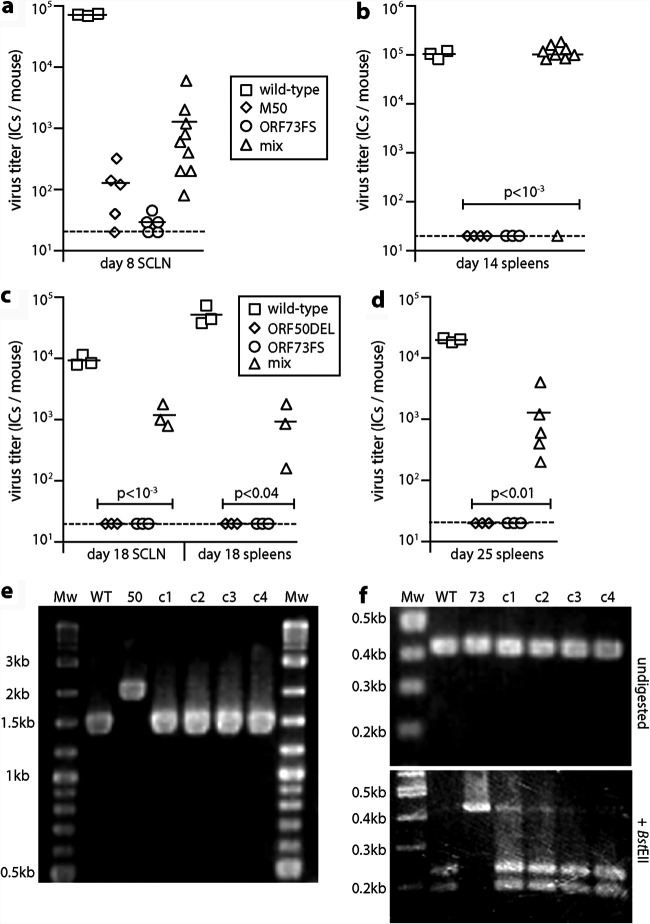
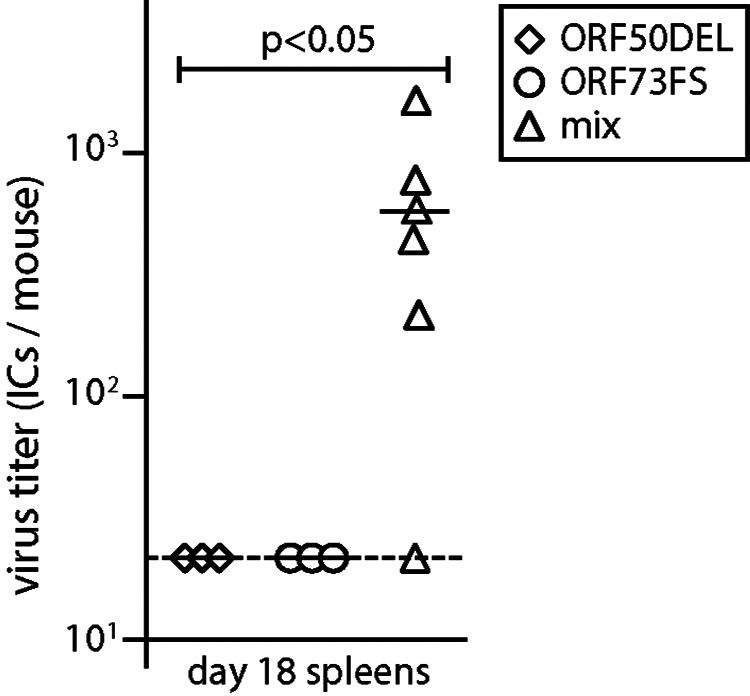
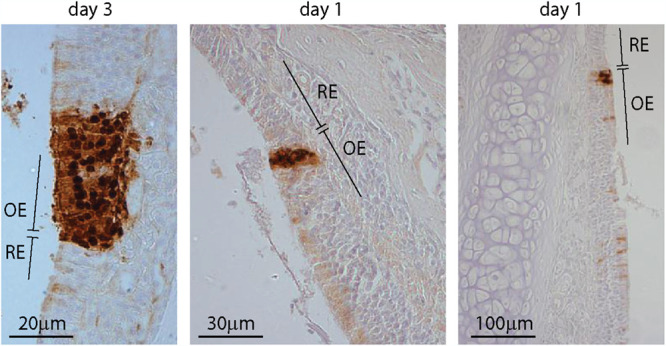
Herpesvirus genomes show abundant evidence of past recombination. Its functional importance is unknown. A key question is whether recombinant viruses can outpace the immunity induced by their parents to reach higher loads. We tested this by coinfecting mice with attenuated mutants of murid herpesvirus 4 (MuHV-4). Infection by the natural olfactory route routinely allowed mutant viruses to reconstitute wild-type genotypes and reach normal viral loads. Lung coinfections rescued much less well. Attenuated murine cytomegalovirus mutants similarly showed recombinational rescue via the nose but not the lungs. These infections spread similarly, so route-specific rescue implied that recombination occurred close to the olfactory entry site. Rescue of replication-deficient MuHV-4 confirmed this, showing that coinfection occurred in the first encountered olfactory cells. This worked even with asynchronous inoculation, implying that a defective virus can wait here for later rescue. Virions entering the nose get caught on respiratory mucus, which the respiratory epithelial cilia push back toward the olfactory surface. Early infection was correspondingly focused on the anterior olfactory edge. Thus, by concentrating incoming infection into a small area, olfactory entry seems to promote functionally significant recombination. IMPORTANCE All organisms depend on genetic diversity to cope with environmental change. Small viruses rely on frequent point mutations. This is harder for herpesviruses because they have larger genomes. Recombination provides another means of genetic optimization. Human herpesviruses often coinfect, and they show evidence of past recombination, but whether this is rare and incidental or functionally important is unknown. We showed that herpesviruses entering mice via the natural olfactory route meet reliably enough for recombination routinely to repair crippling mutations and restore normal viral loads. It appeared to occur in the first encountered olfactory cells and reflected a concentration of infection at the anterior olfactory edge. Thus, natural host entry incorporates a significant capacity for herpesvirus recombination.
Keywords: herpesviruses; host entry; olfactory; recombination.
Figures
References
-
- Suárez NM, Wilkie GS, Hage E, Camiolo S, Holton M, Hughes J, Maabar M, Vattipally SB, Dhingra A, Gompels UA, Wilkinson GWG, Baldanti F, Furione M, Lilleri D, Arossa A, Ganzenmueller T, Gerna G, Hubáček P, Schulz TF, Wolf D, Zavattoni M, Davison AJ. 2019. Human cytomegalovirus genomes sequenced directly from clinical material: variation, multiple-strain infection, recombination, and gene loss. J Infect Dis 220:781–791. 10.1093/infdis/jiz208. - DOI - PMC - PubMed
-
- Palser AL, Grayson NE, White RE, Corton C, Correia S, Ba Abdullah MM, Watson SJ, Cotten M, Arrand JR, Murray PG, Allday MJ, Rickinson AB, Young LS, Farrell PJ, Kellam P. 2015. Genome diversity of Epstein-Barr virus from multiple tumor types and normal infection. J Virol 89:5222–5237. 10.1128/JVI.03614-14. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
