

Nanoscale molecular architecture controls calcium diffusion and ER replenishment in dendritic spines
- PMID: 34524854
- PMCID: PMC8443180
- DOI: 10.1126/sciadv.abh1376
Nanoscale molecular architecture controls calcium diffusion and ER replenishment in dendritic spines
Abstract
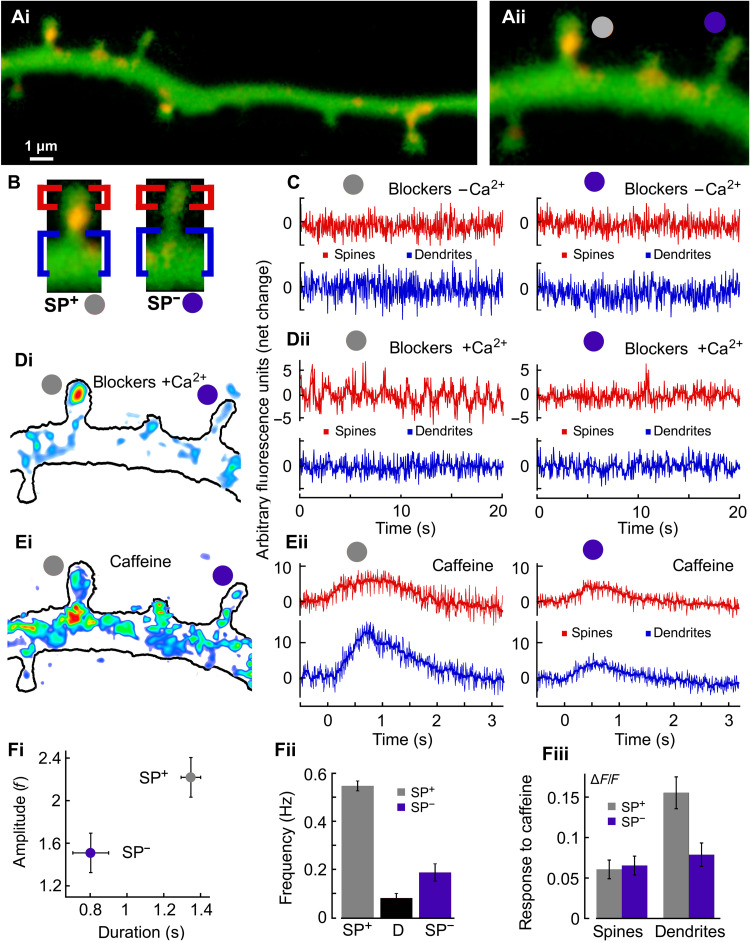
Dendritic spines are critical components of neuronal synapses as they receive and transform synaptic inputs into a succession of calcium-regulated biochemical events. The spine apparatus (SA), an extension of smooth endoplasmic reticulum, regulates slow and fast calcium dynamics in spines. Calcium release events deplete SA calcium ion reservoir rapidly, yet the next cycle of signaling requires its replenishment. How spines achieve this replenishment without triggering calcium release remains unclear. Using computational modeling, calcium and STED superresolution imaging, we show that the SA replenishment involves the store-operated calcium entry pathway during spontaneous calcium transients. We identified two main conditions for SA replenishment without depletion: a small amplitude and a slow timescale for calcium influx, and a close proximity between SA and plasma membranes. Thereby, spine’s nanoscale organization separates SA replenishment from depletion. We further conclude that spine’s receptor organization also determines the calcium dynamics during the induction of long-term synaptic changes.
Figures





References
-
- Cummings J. A., Mulkey R. M., Nicoll R. A., Malenka R. C., Ca2+ signaling requirements for long-term depression in the hippocampus. Neuron 16, 825–833 (1996). - PubMed
-
- Segal M., Dendritic spines and long-term plasticity. Nat. Rev. Neurosci. 6, 277–284 (2005). - PubMed
-
- R. Yuste, Dendritic Spines (MIT press, 2010).
-
- Yuste R., Bonhoeffer T., Morphological changes in dendritic spines associated with long-term synaptic plasticity. Annu. Rev. Neurosci. 24, 1071–1089 (2001). - PubMed
LinkOut - more resources
Full Text Sources
