

Resident memory CD8+ T cells in regional lymph nodes mediate immunity to metastatic melanoma
- PMID: 34525340
- PMCID: PMC9015193
- DOI: 10.1016/j.immuni.2021.08.019
Resident memory CD8+ T cells in regional lymph nodes mediate immunity to metastatic melanoma
Abstract
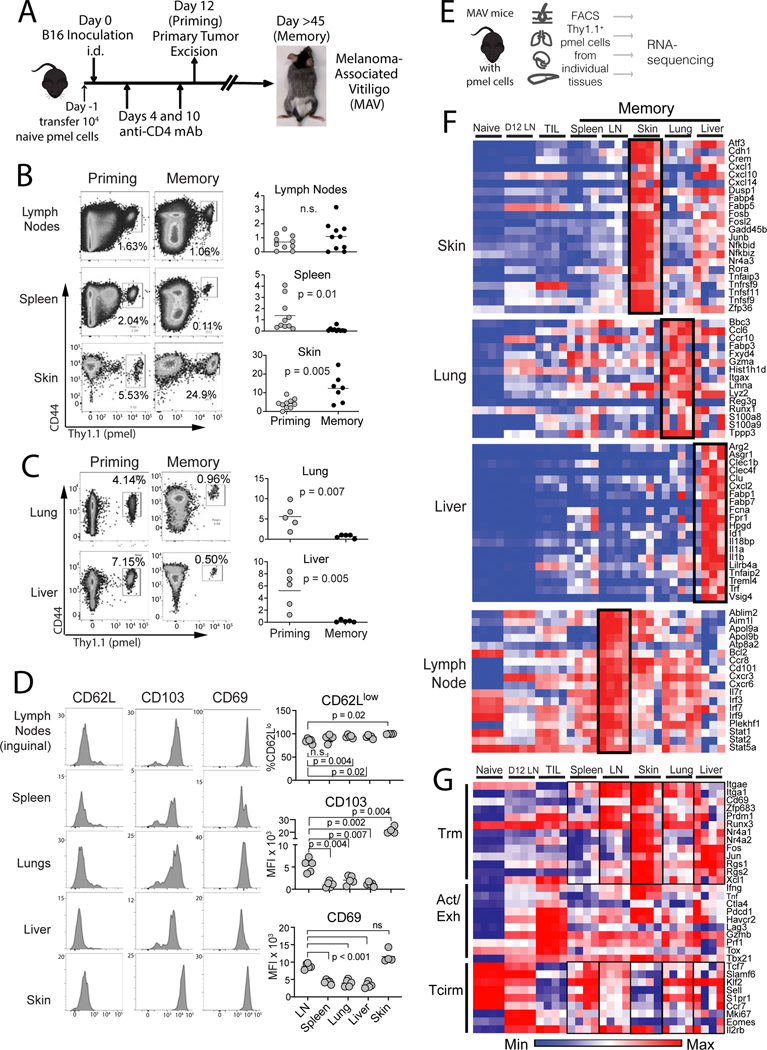
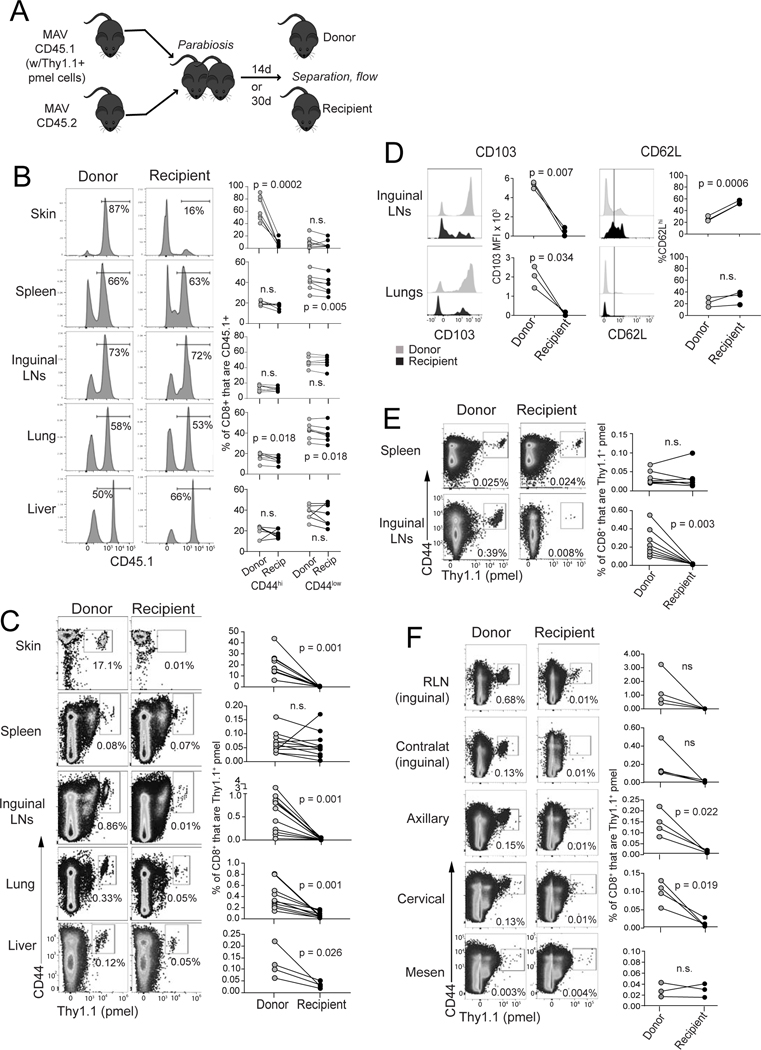
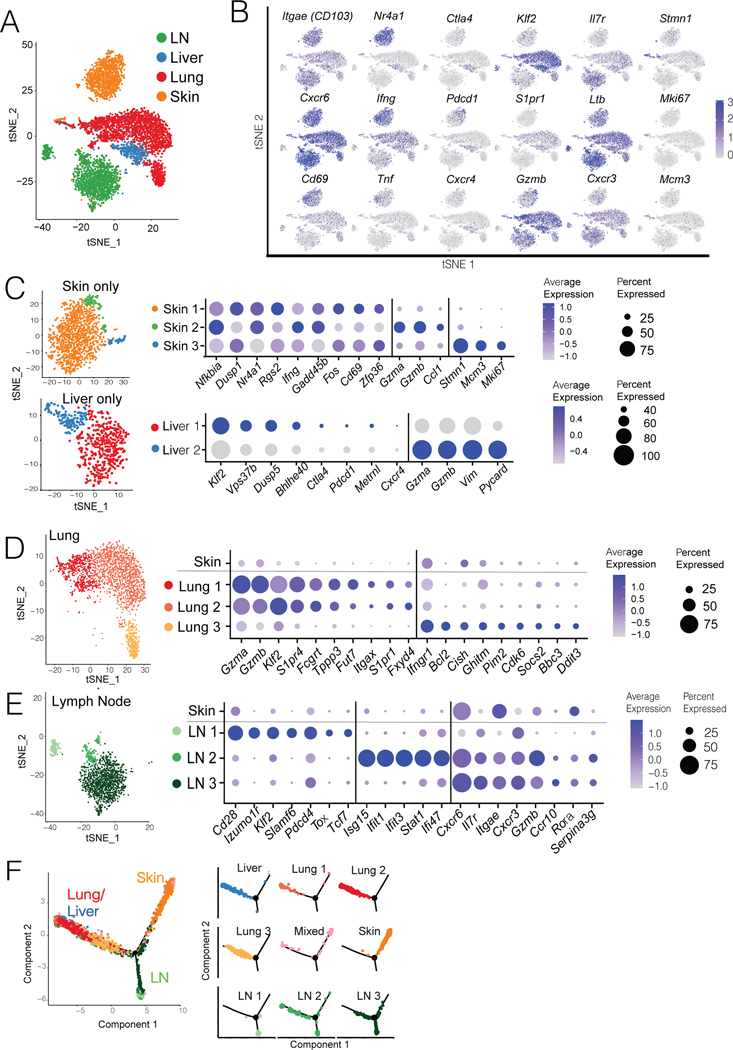
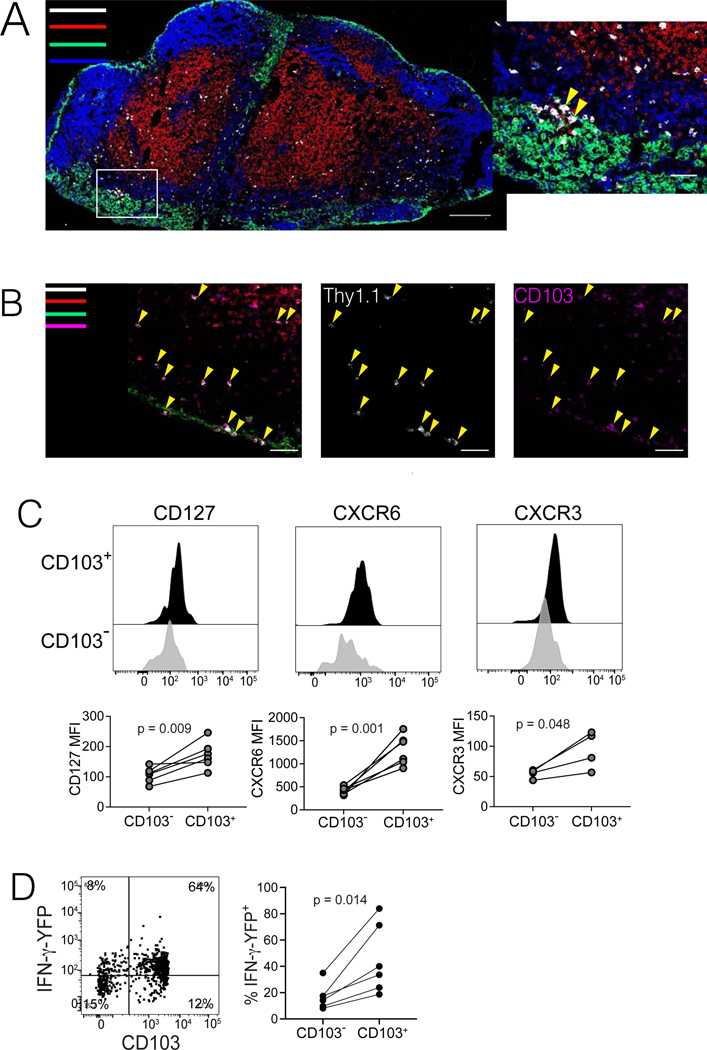
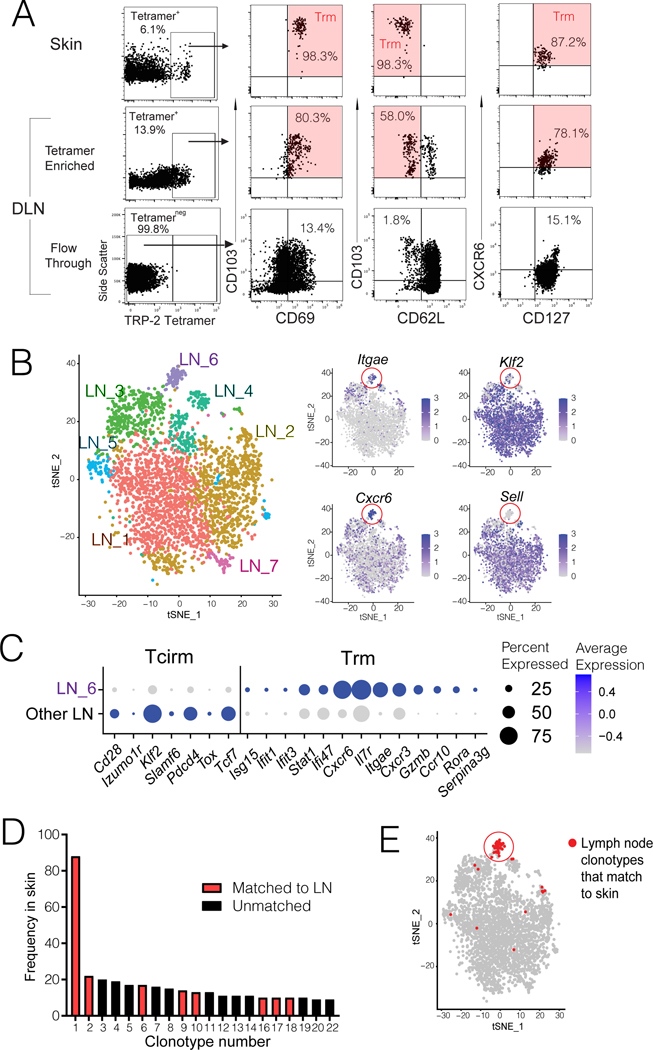
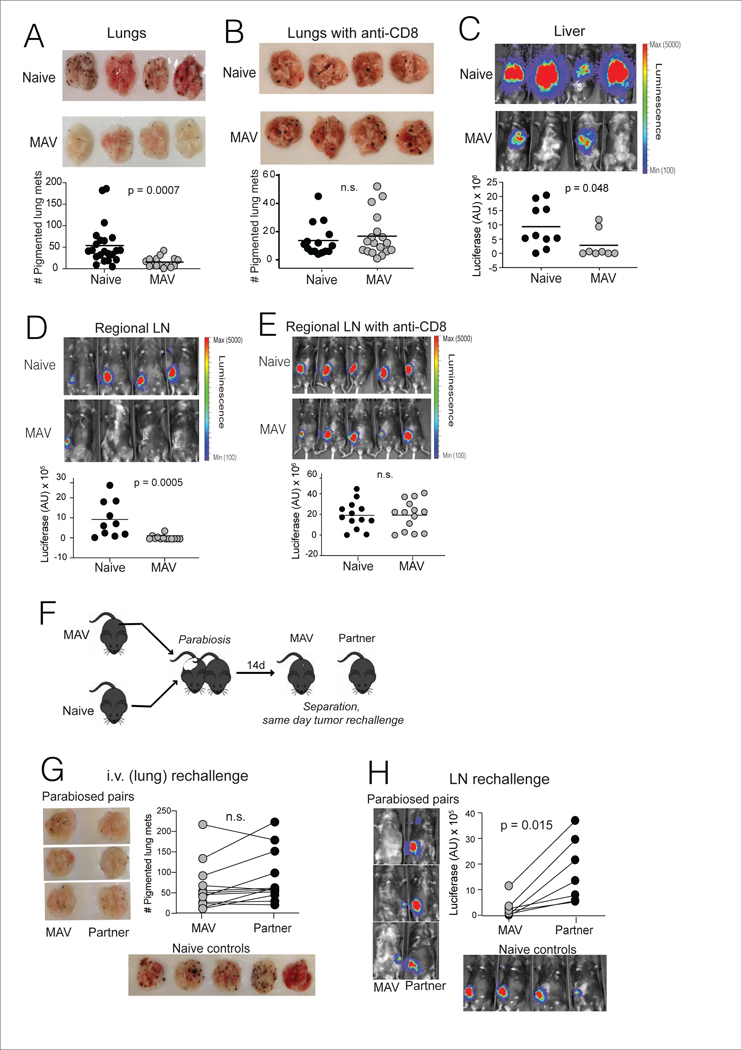
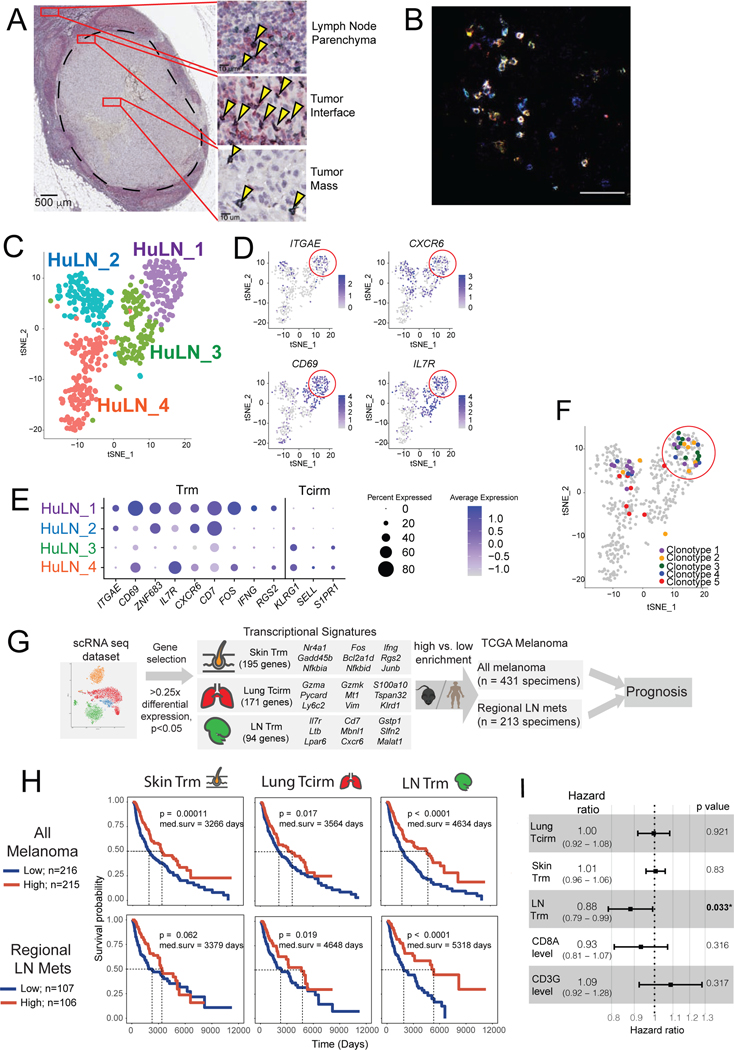
The nature of the anti-tumor immune response changes as primary tumors progress and metastasize. We investigated the role of resident memory (Trm) and circulating memory (Tcirm) cells in anti-tumor responses at metastatic locations using a mouse model of melanoma-associated vitiligo. We found that the transcriptional characteristics of tumor-specific CD8+ T cells were defined by the tissue of occupancy. Parabiosis revealed that tumor-specific Trm and Tcirm compartments persisted throughout visceral organs, but Trm cells dominated lymph nodes (LNs). Single-cell RNA-sequencing profiles of Trm cells in LN and skin were distinct, and T cell clonotypes that occupied both tissues were overwhelmingly maintained as Trm in LNs. Whereas Tcirm cells prevented melanoma growth in the lungs, Trm afforded long-lived protection against melanoma seeding in LNs. Expanded Trm populations were also present in melanoma-involved LNs from patients, and their transcriptional signature predicted better survival. Thus, tumor-specific Trm cells persist in LNs, restricting metastatic cancer.
Keywords: CD69; CD8 T cells; CXCR6; Cancer; TCR; TRP-2; Trm; parabiosis; scRNA-seq; vitiligo.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests Authors declare no competing interests.
Figures
References
-
- Asano K, Nabeyama A, Miyake Y, Qiu CH, Kurita A, Tomura M, Kanagawa O, Fujii S, and Tanaka M. (2011). CD169-positive macrophages dominate antitumor immunity by crosspresenting dead cell-associated antigens. Immunity 34, 85–95. - PubMed
-
- Brown M, Assen FP, Leithner A, Abe J, Schachner H, Asfour G, Bago-Horvath Z, Stein JV, Uhrin P, Sixt M, and Kerjaschki D. (2018). Lymph node blood vessels provide exit routes for metastatic tumor cell dissemination in mice. Science 359, 1408–1411. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
