

Extrusion of chromatin loops by a composite loop extrusion factor
- PMID: 34525654
- PMCID: PMC9112126
- DOI: 10.1103/PhysRevE.104.024414
Extrusion of chromatin loops by a composite loop extrusion factor
Abstract
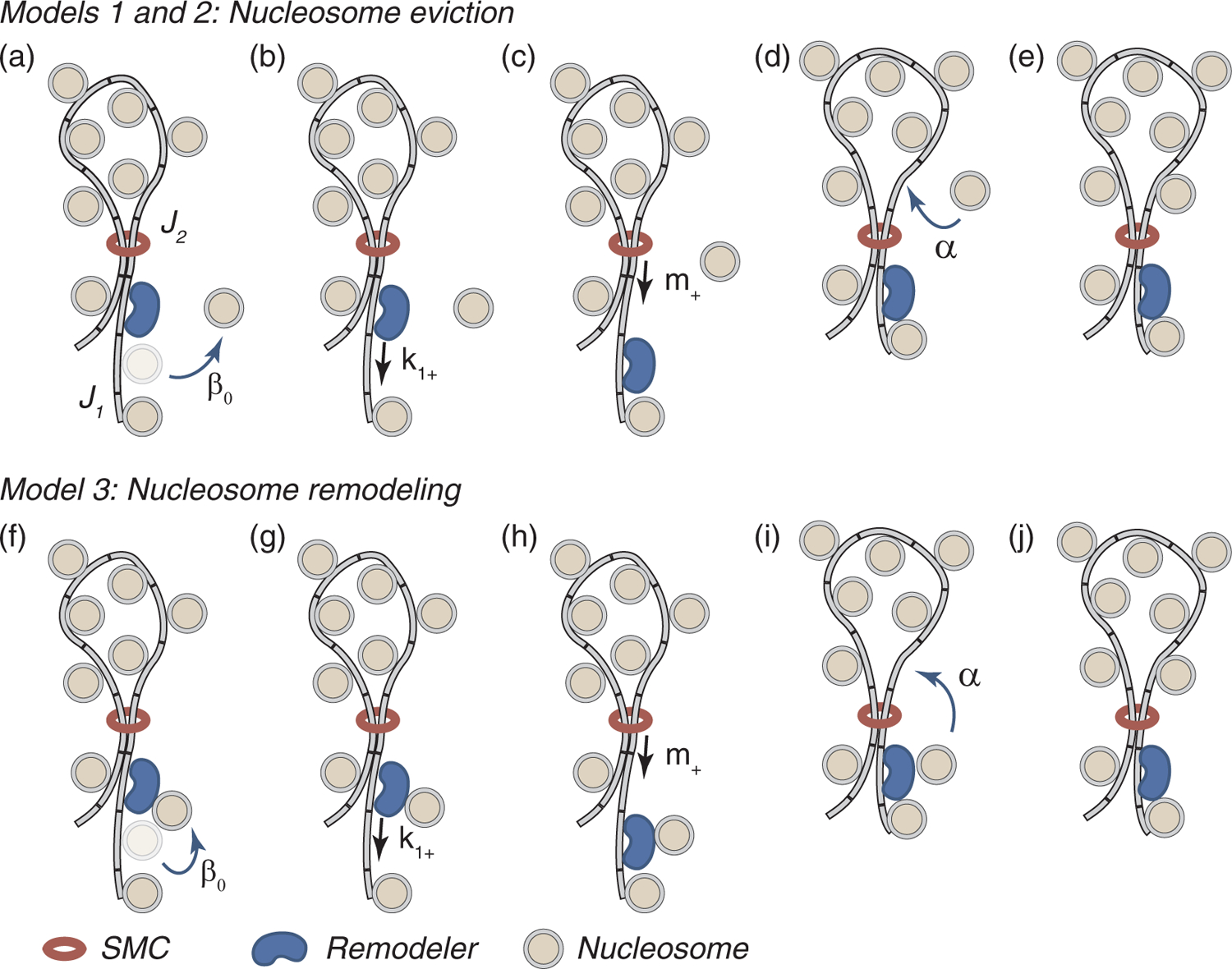
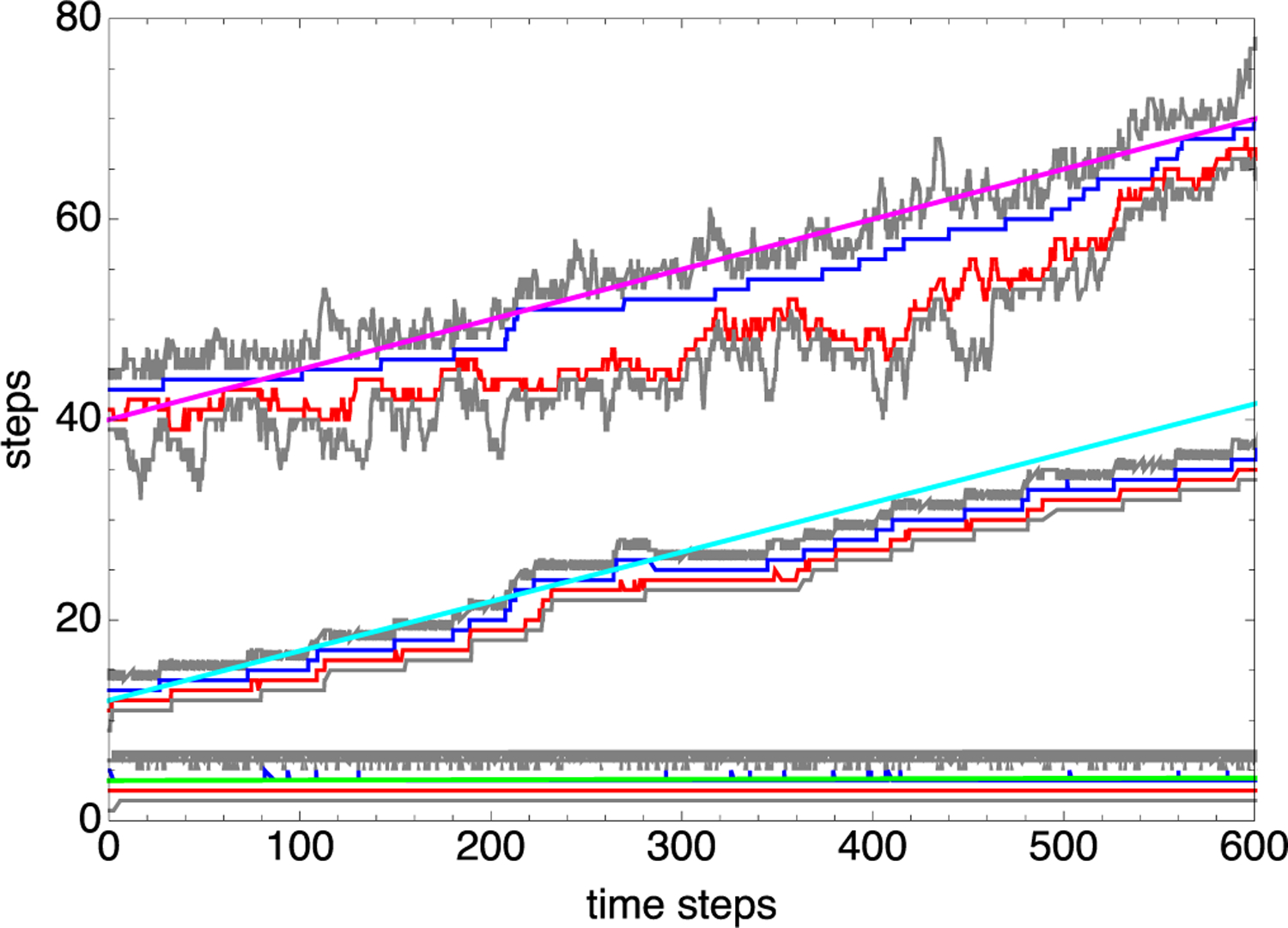
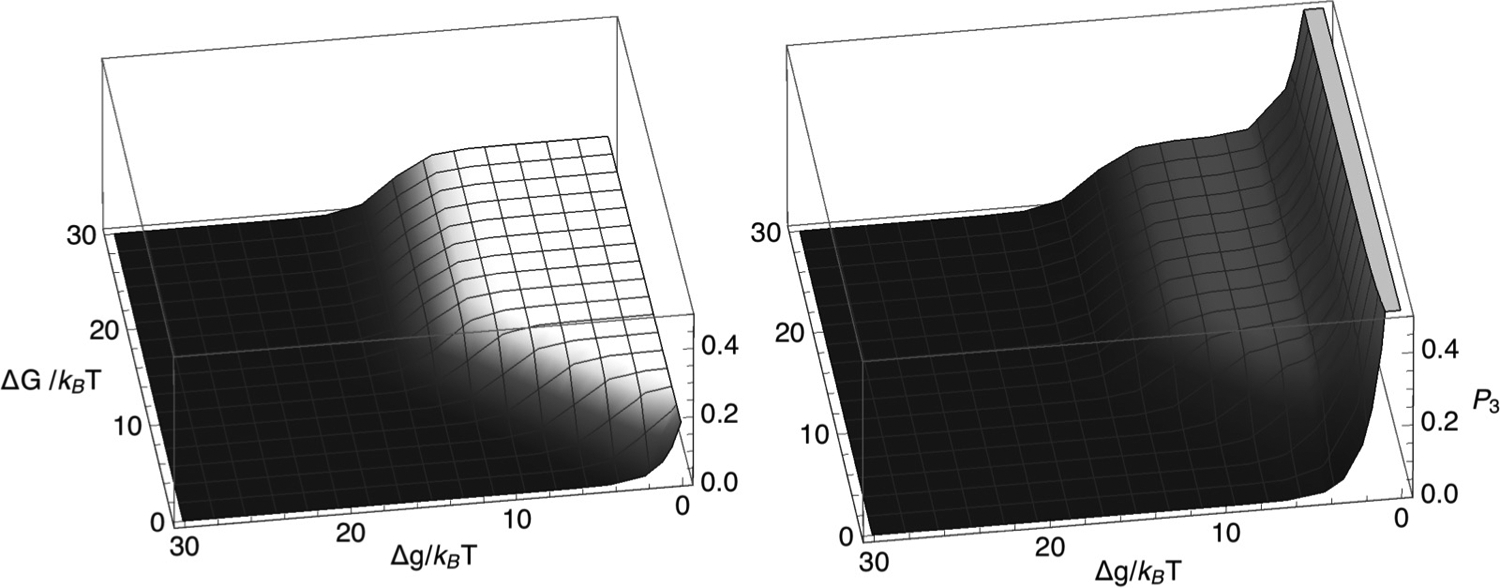
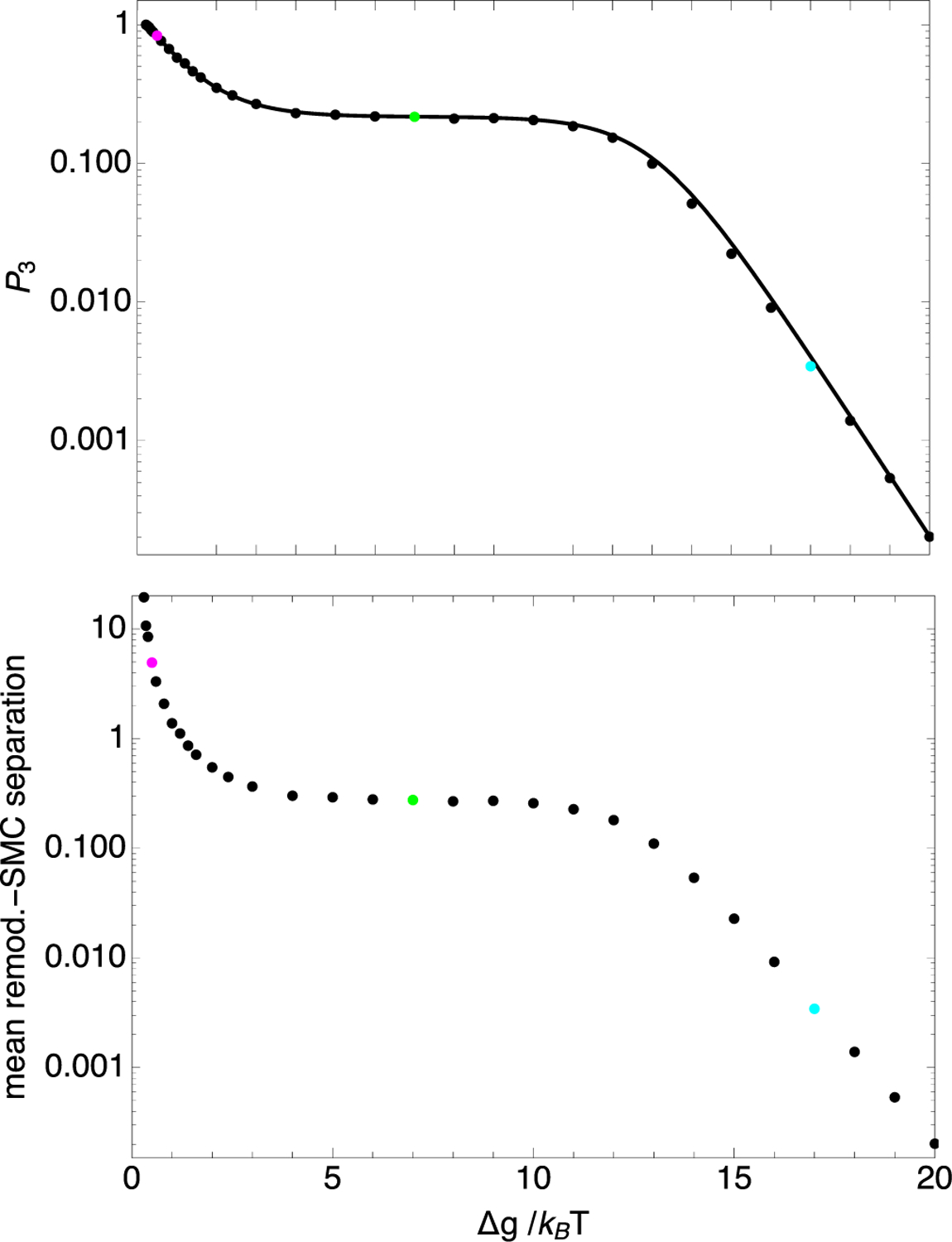
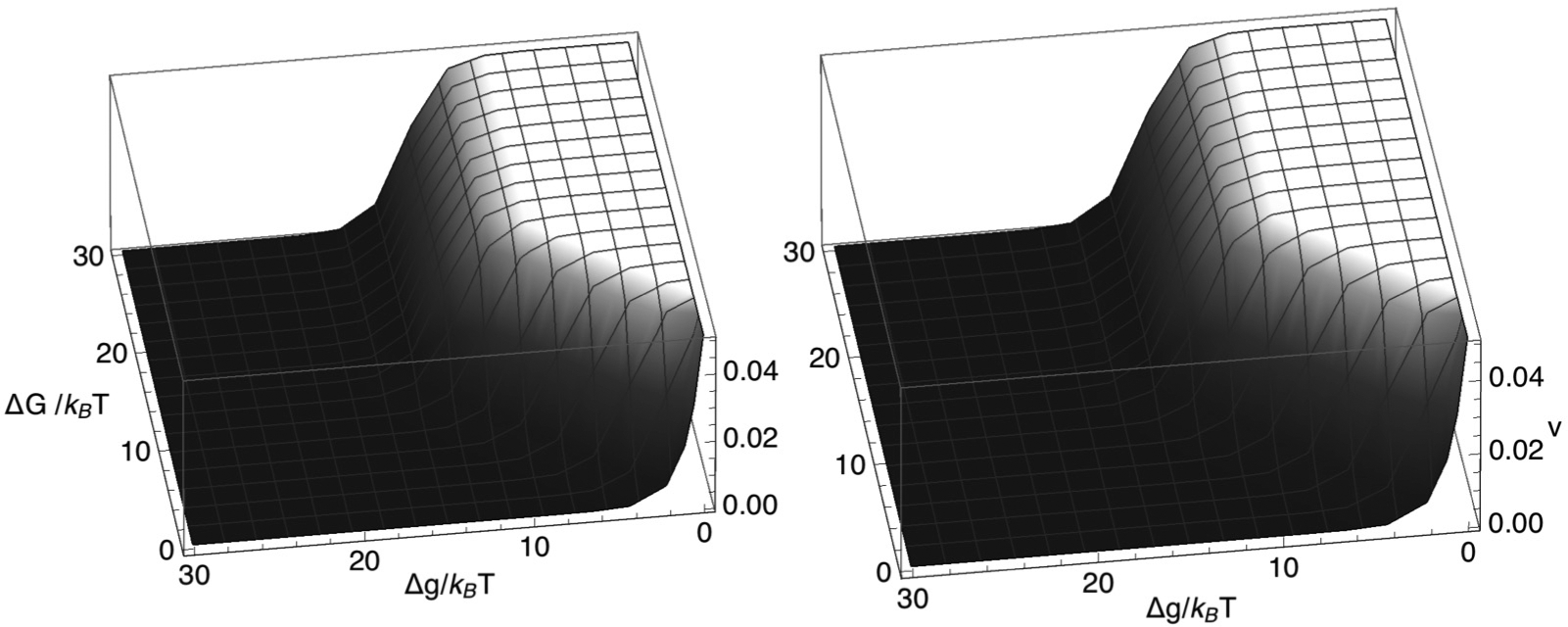
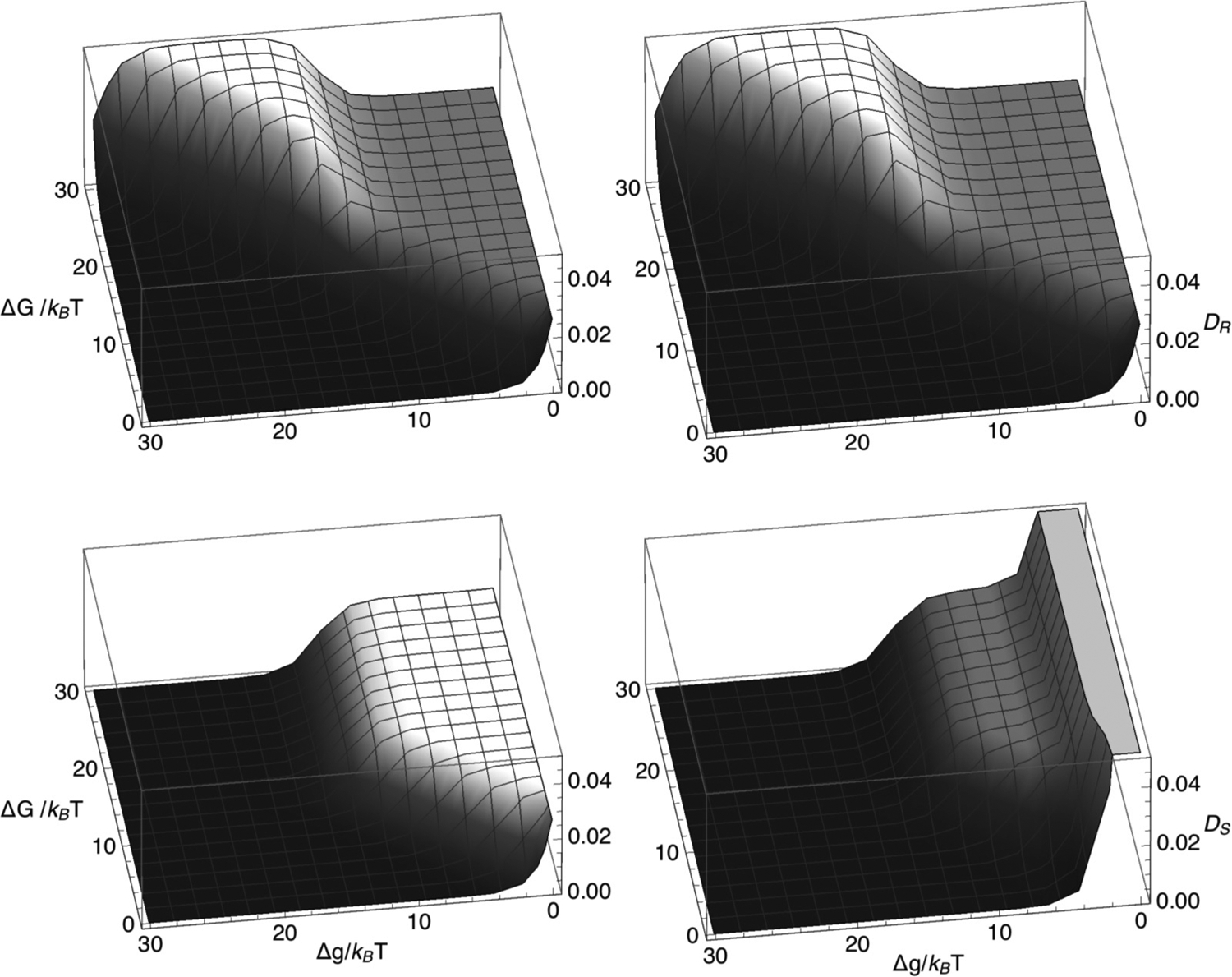
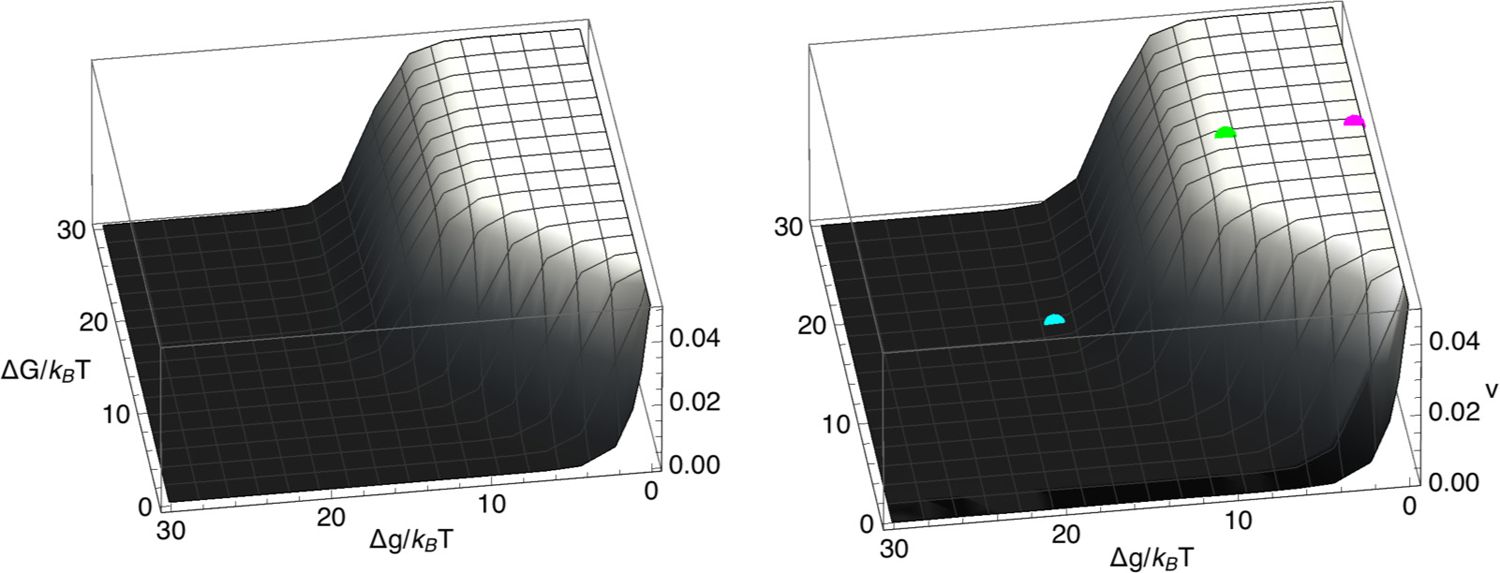
Chromatin loop extrusion by structural maintenance of chromosome (SMC) complexes is thought to underlie intermediate-scale chromatin organization inside cells. Motivated by a number of experiments suggesting that nucleosomes may block loop extrusion by SMCs, such as cohesin and condensin complexes, we introduce and characterize theoretically a composite loop extrusion factor (composite LEF) model. In addition to an SMC complex that creates a chromatin loop by encircling two threads of DNA, this model includes a remodeling complex that relocates or removes nucleosomes as it progresses along the chromatin, and nucleosomes that block SMC translocation along the DNA. Loop extrusion is enabled by SMC motion along nucleosome-free DNA, created in the wake of the remodeling complex, while nucleosome rebinding behind the SMC acts as a ratchet, holding the SMC close to the remodeling complex. We show that, for a wide range of parameter values, this collection of factors constitutes a composite LEF that extrudes loops with a velocity, comparable to the velocity of remodeling complex translocation on chromatin in the absence of SMC, and much faster than loop extrusion by an isolated SMC that is blocked by nucleosomes.
Figures
References
-
- Dekker J, Rippe K, Dekker M, and Kleckner N, Capturing chromosome conformation, Science 295, 1306 (2002). - PubMed
-
- Sexton T, Yaffe E, Kenigsberg E, Bantignies F, Leblanc B, Hoichman M, Parrinello H, Tanay A, and Cavalli G, Three-dimensional folding and functional organization principles of the Drosophila genome, Cell 148, 458 (2012). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources