

p38 MAPK-mediated loss of nuclear RNase III enzyme Drosha underlies amyloid beta-induced neuronal stress in Alzheimer's disease
- PMID: 34528746
- PMCID: PMC8521488
- DOI: 10.1111/acel.13434
p38 MAPK-mediated loss of nuclear RNase III enzyme Drosha underlies amyloid beta-induced neuronal stress in Alzheimer's disease
Abstract
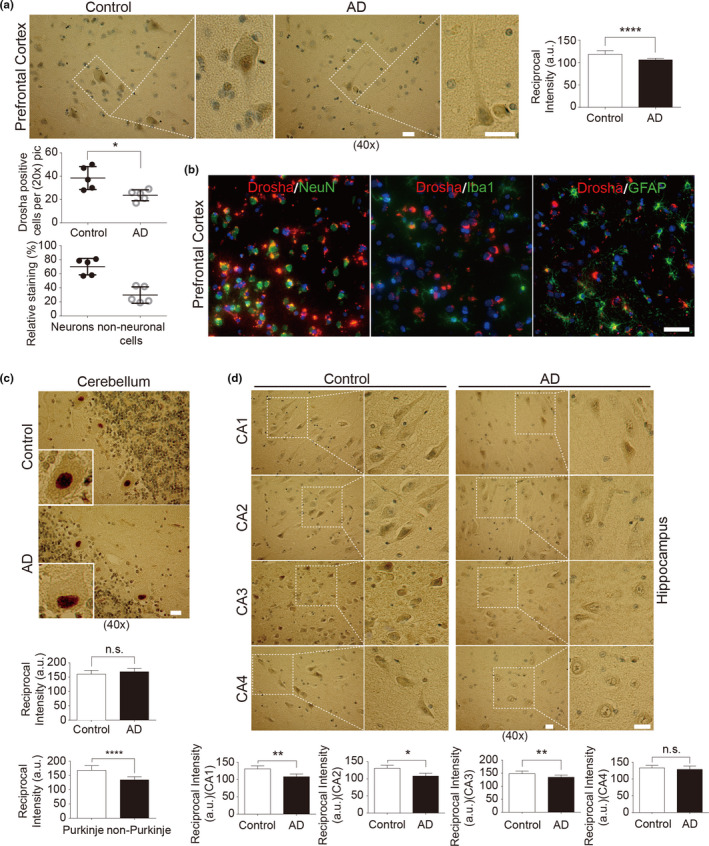
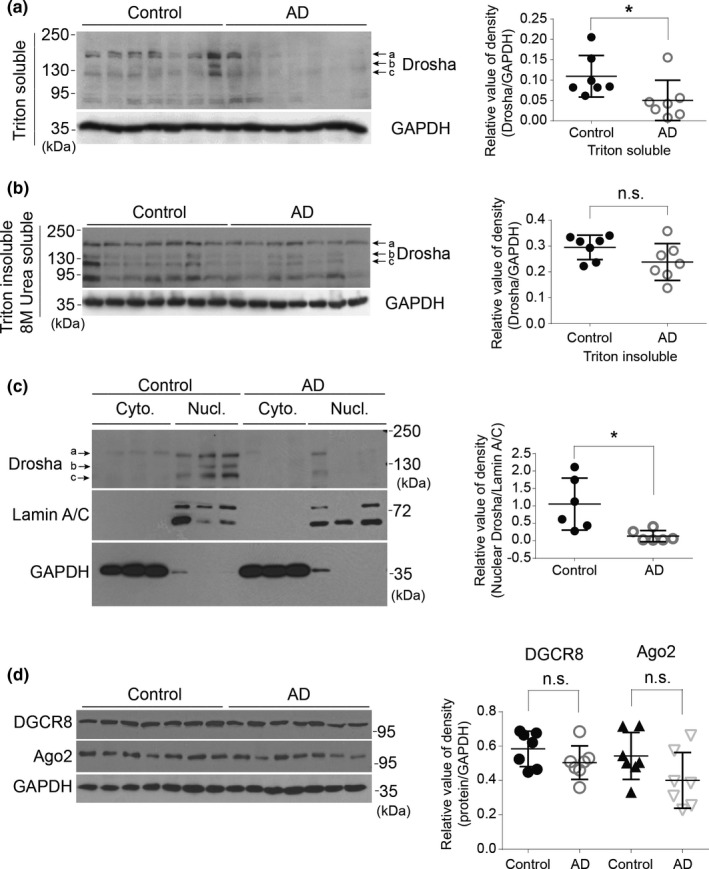
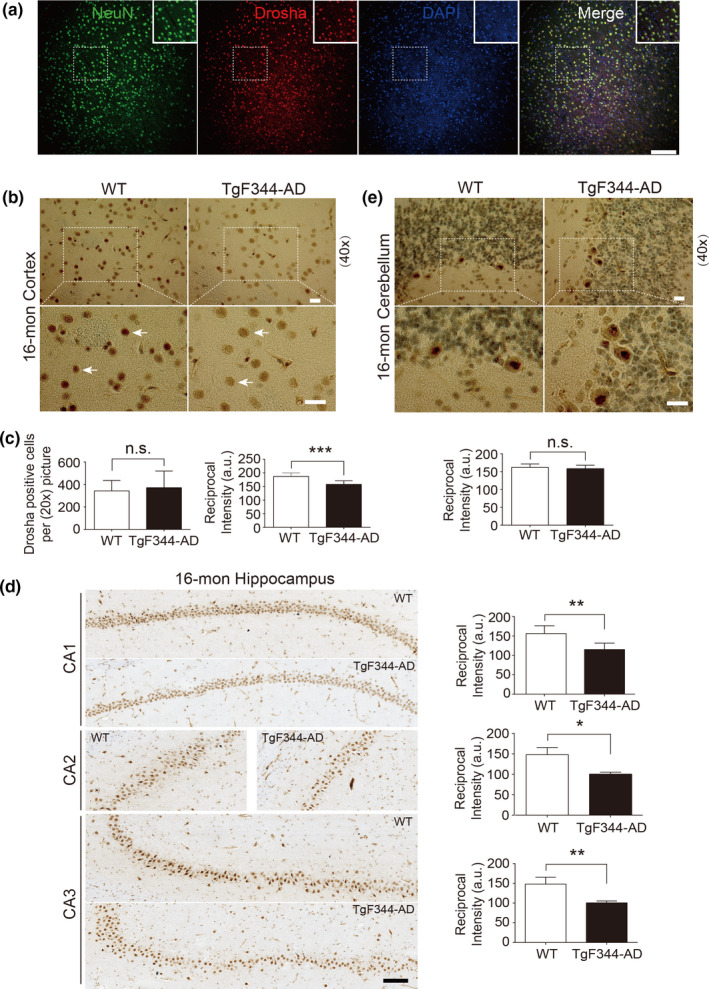
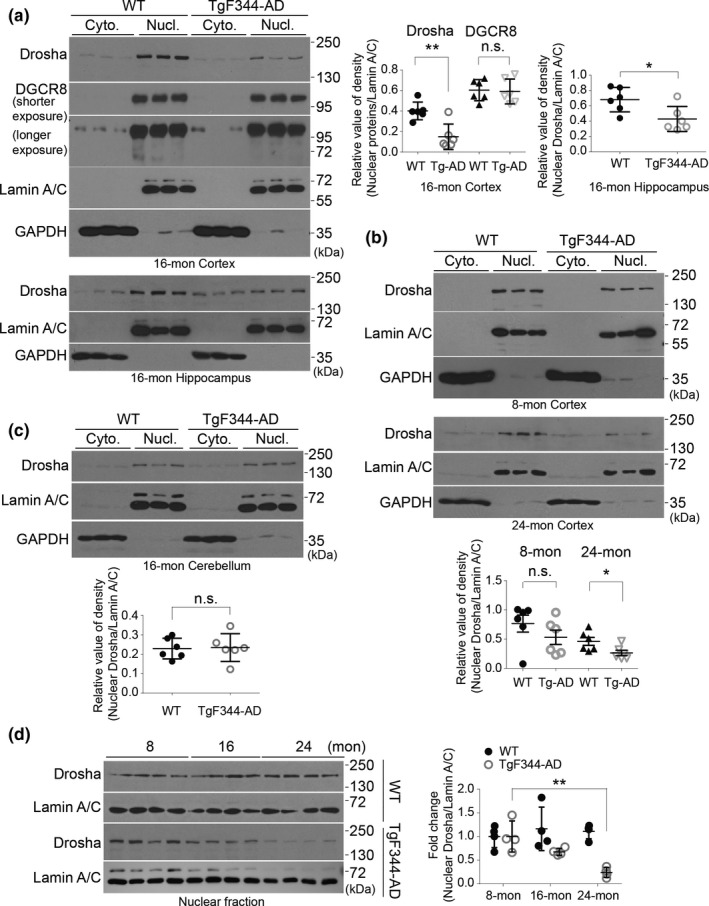
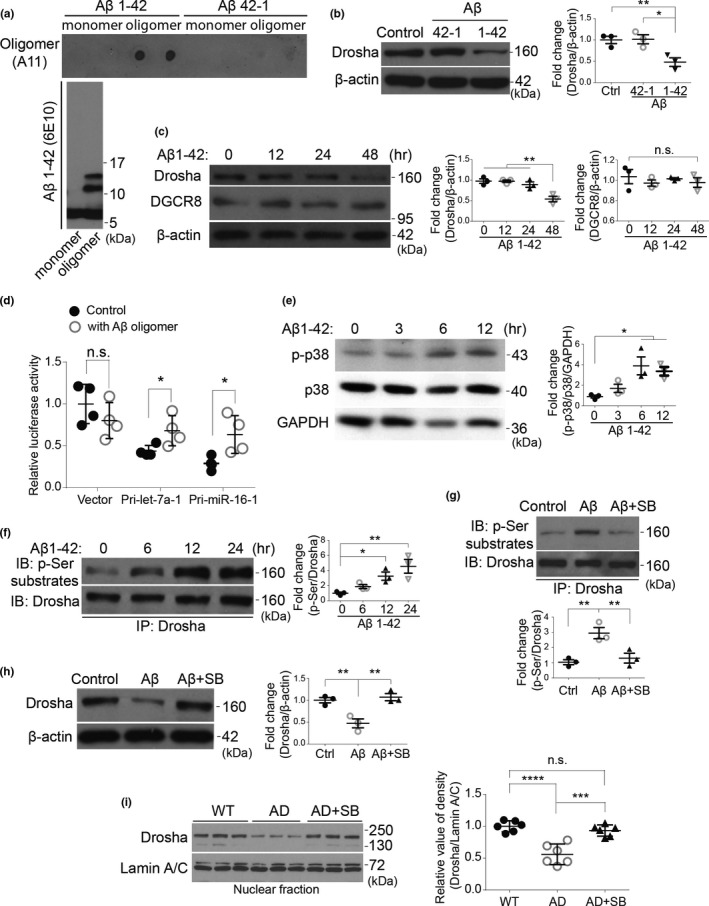
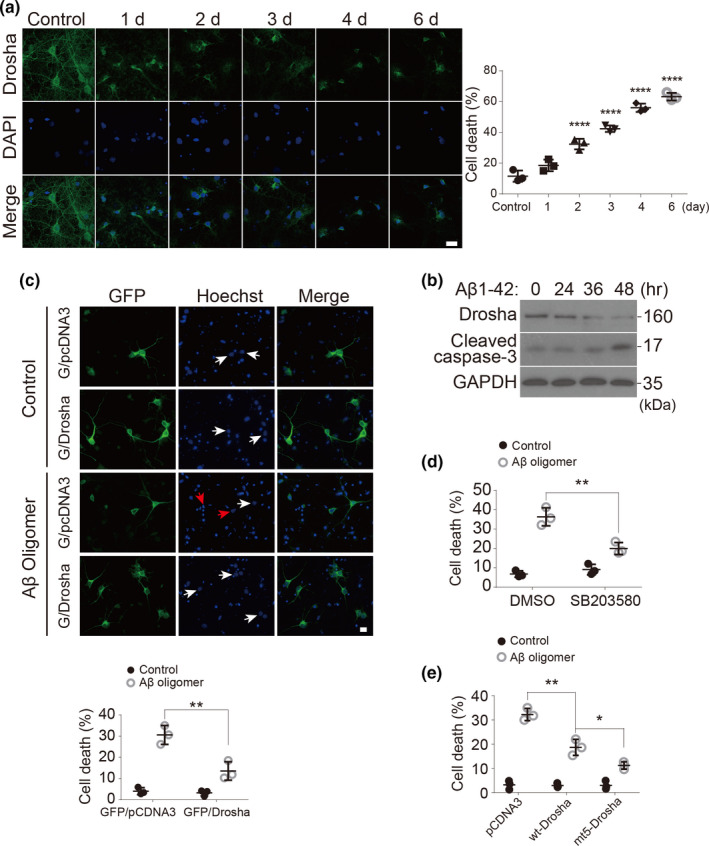
MicroRNAs (miRNAs) are small noncoding RNAs ubiquitously expressed in the brain and regulate gene expression at the post-transcriptional level. The nuclear RNase III enzyme Drosha initiates the maturation process of miRNAs in the nucleus. Strong evidence suggests that dysregulation of miRNAs is involved in many neurological disorders including Alzheimer's disease (AD). Dysfunction of miRNA biogenesis components may be involved in the processes of those diseases. However, the role of Drosha in AD remains unknown. By using immunohistochemistry, biochemistry, and subcellular fractionation methods, we show here that the level of Drosha protein was significantly lower in the postmortem brain of human AD patients as well as in the transgenic rat model of AD. Interestingly, Drosha level was specifically reduced in neurons of the cortex and hippocampus but not in the cerebellum in the AD brain samples. In primary cortical neurons, amyloid-beta (Aβ) oligomers caused a p38 MAPK-dependent phosphorylation of Drosha, leading to its redistribution from the nucleus to the cytoplasm and a decrease in its level. This loss of Drosha function preceded Aβ-induced neuronal death. Importantly, inhibition of p38 MAPK activity or overexpression of Drosha protected neurons from Aβ oligomers-induced apoptosis. Taken together, these results establish a role for p38 MAPK-Drosha pathway in modulating neuronal viability under Aβ oligomers stress condition and implicate loss of Drosha as a key molecular change in the pathogenesis of AD.
Keywords: Alzheimer's disease; Drosha; amyloid beta (Aβ); neuronal death; p38 MAPK.
© 2021 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Babiarz, J. E. , Hsu, R. , Melton, C. , Thomas, M. , Ullian, E. M. , & Blelloch, R. (2011). A role for noncanonical microRNAs in the mammalian brain revealed by phenotypic differences in Dgcr8 versus Dicer1 knockouts and small RNA sequencing. RNA, 17(8), 1489–1501. 10.1261/rna.2442211 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
