

Control of topoisomerase II activity and chemotherapeutic inhibition by TCA cycle metabolites
- PMID: 34529934
- PMCID: PMC8913808
- DOI: 10.1016/j.chembiol.2021.08.014
Control of topoisomerase II activity and chemotherapeutic inhibition by TCA cycle metabolites
Abstract
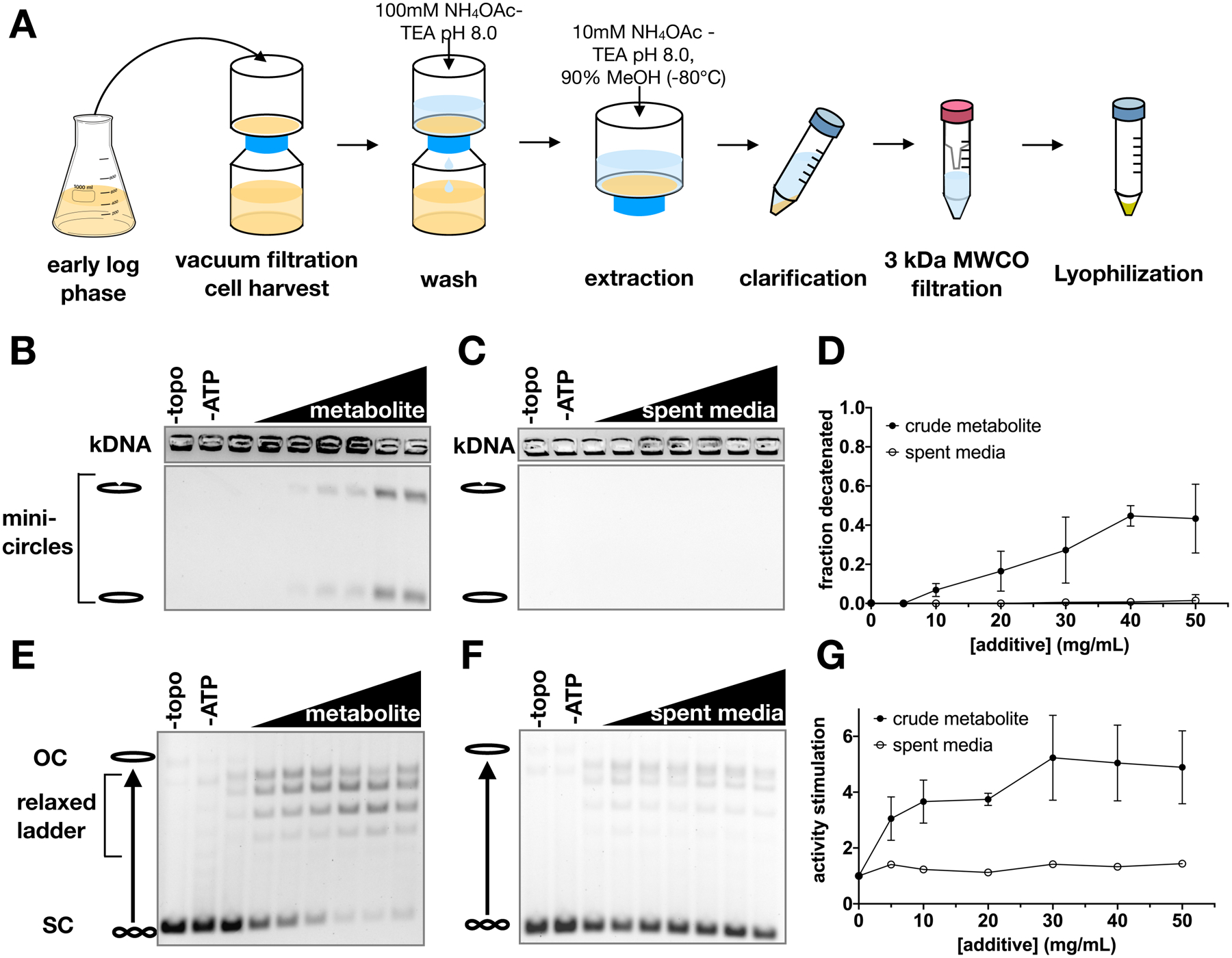
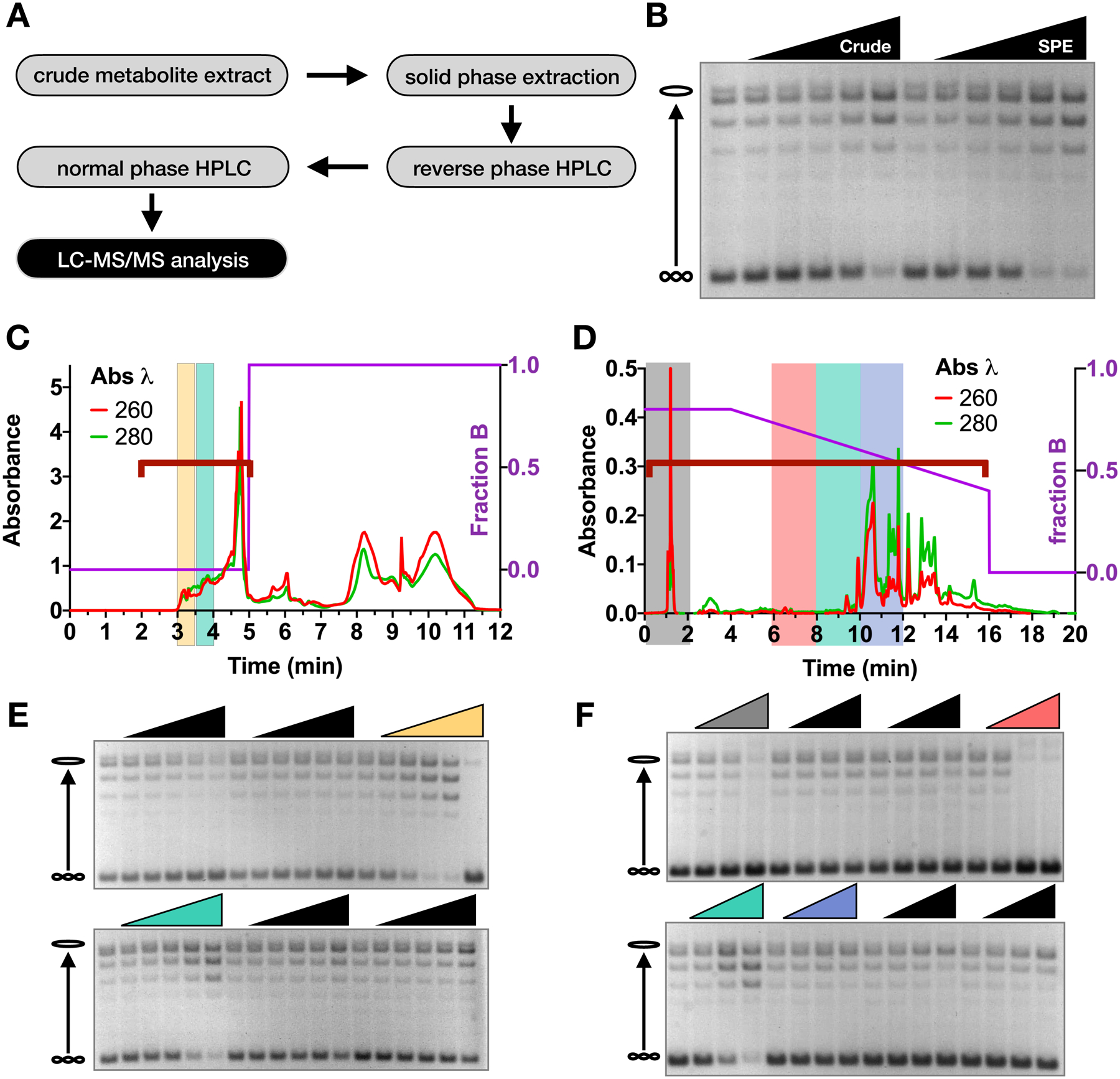
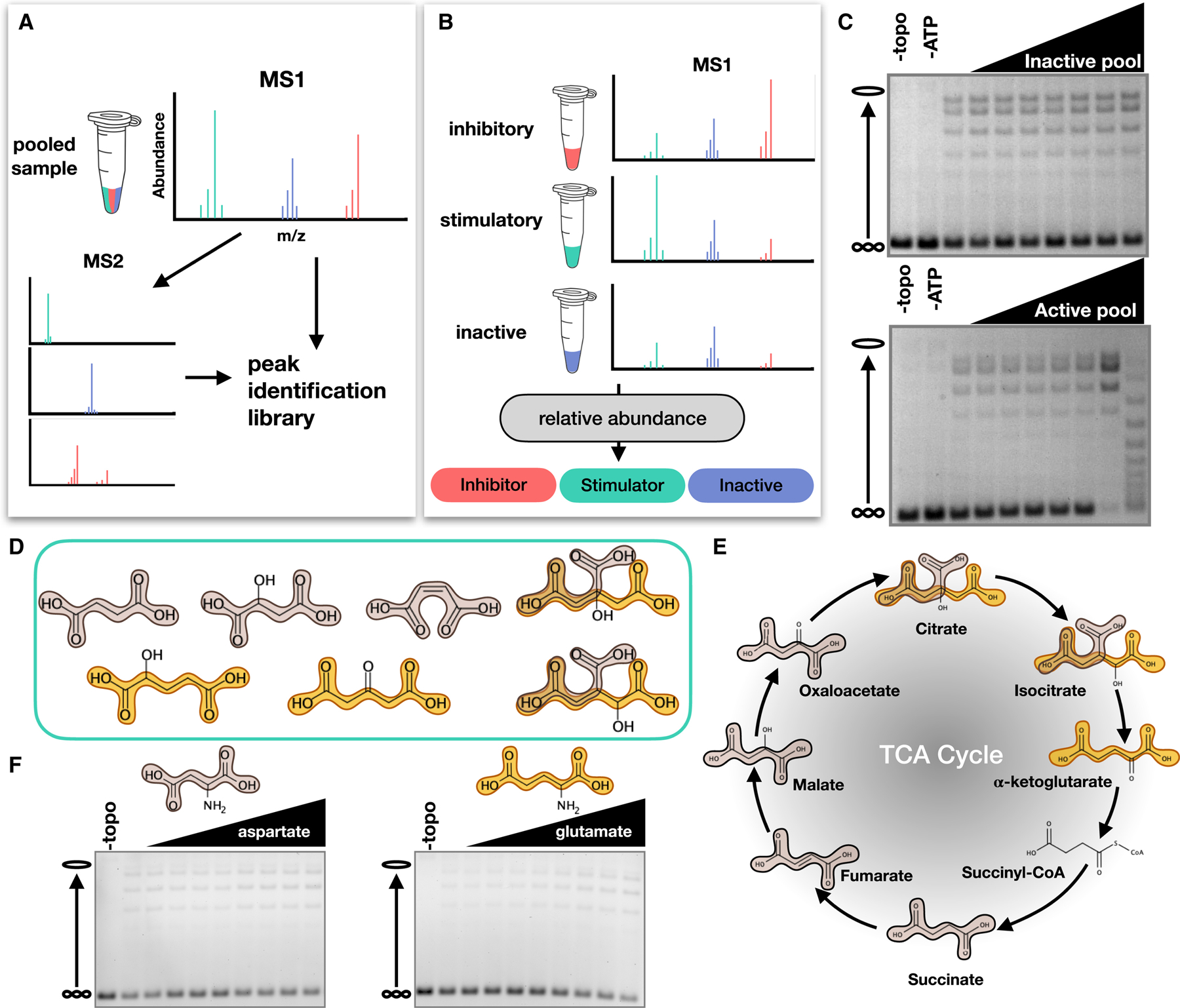
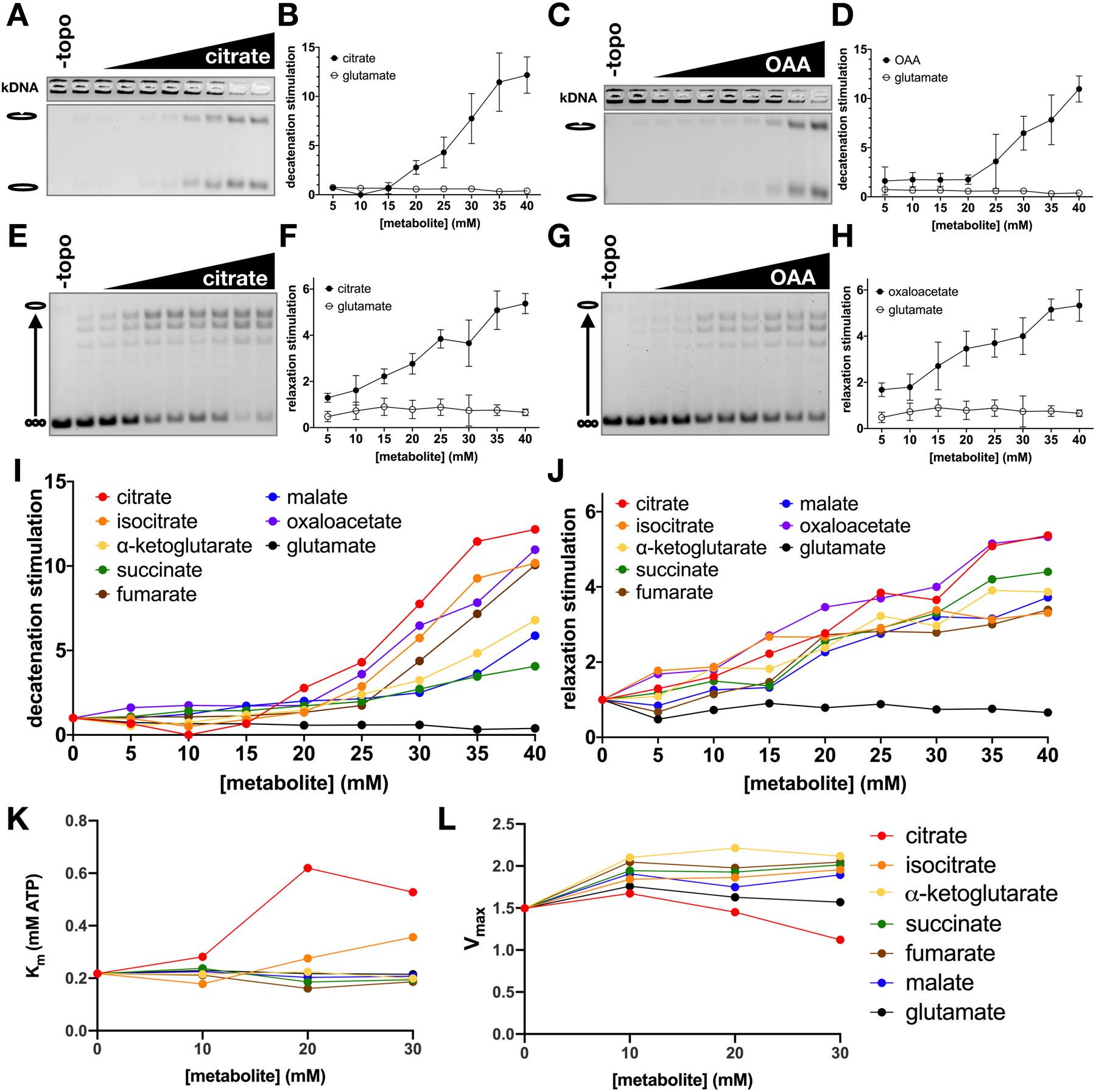
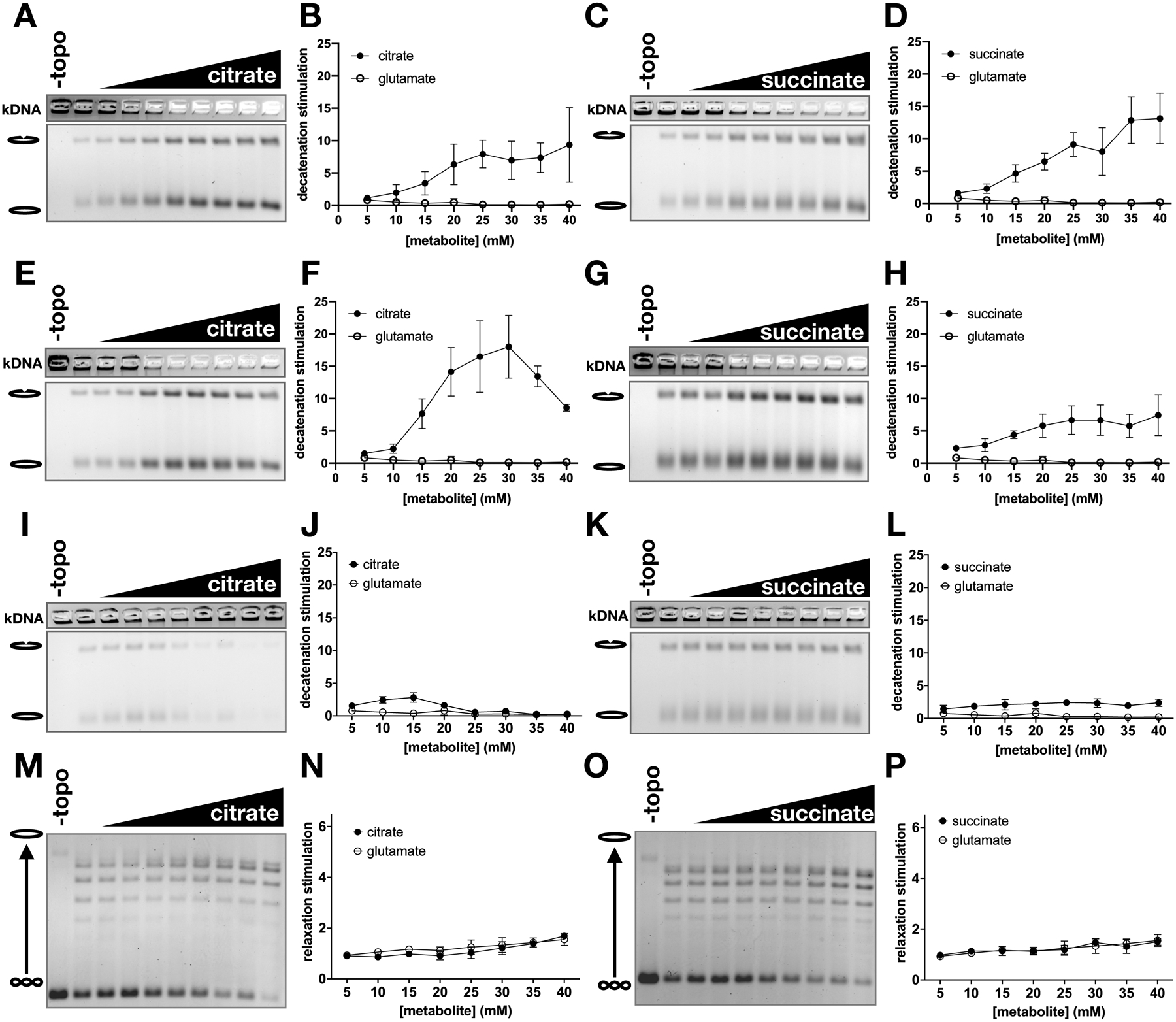
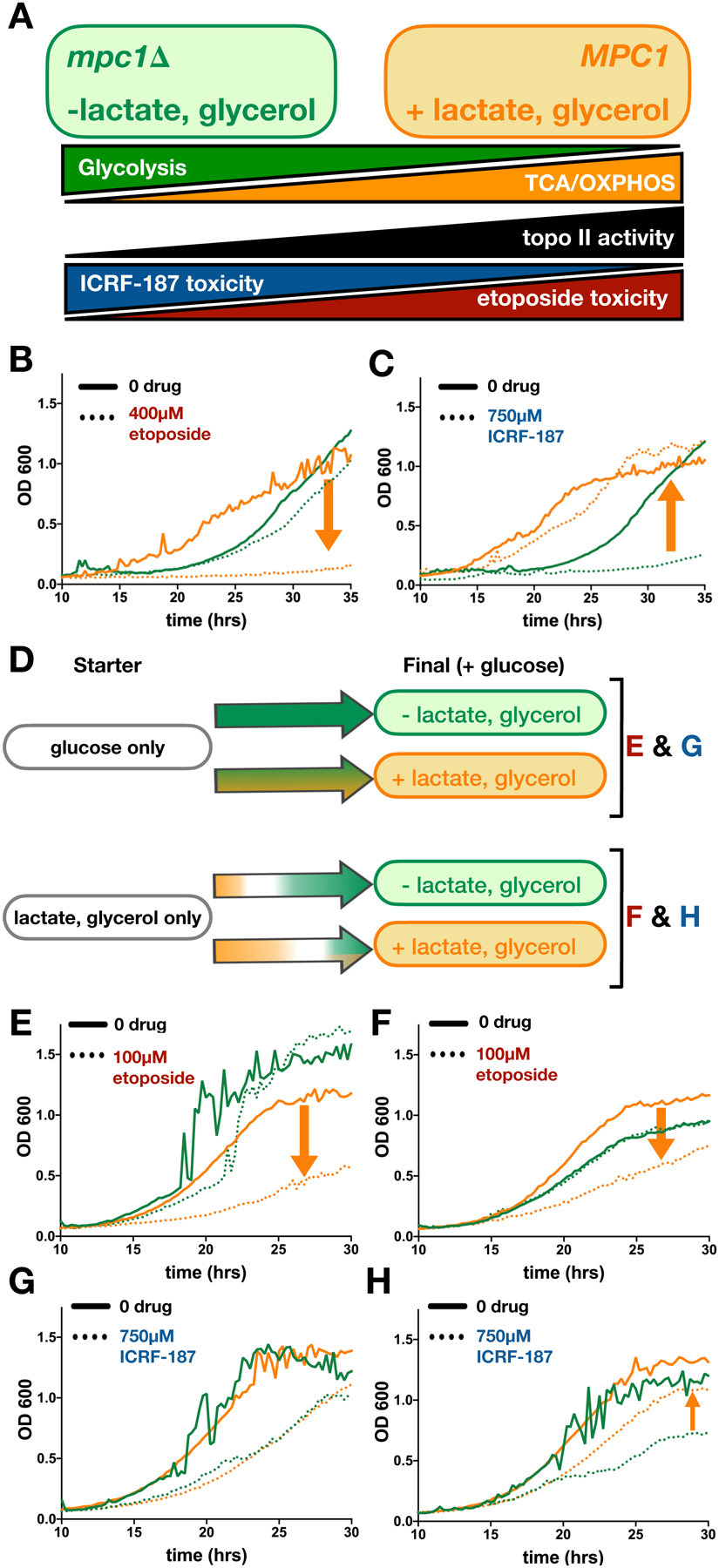
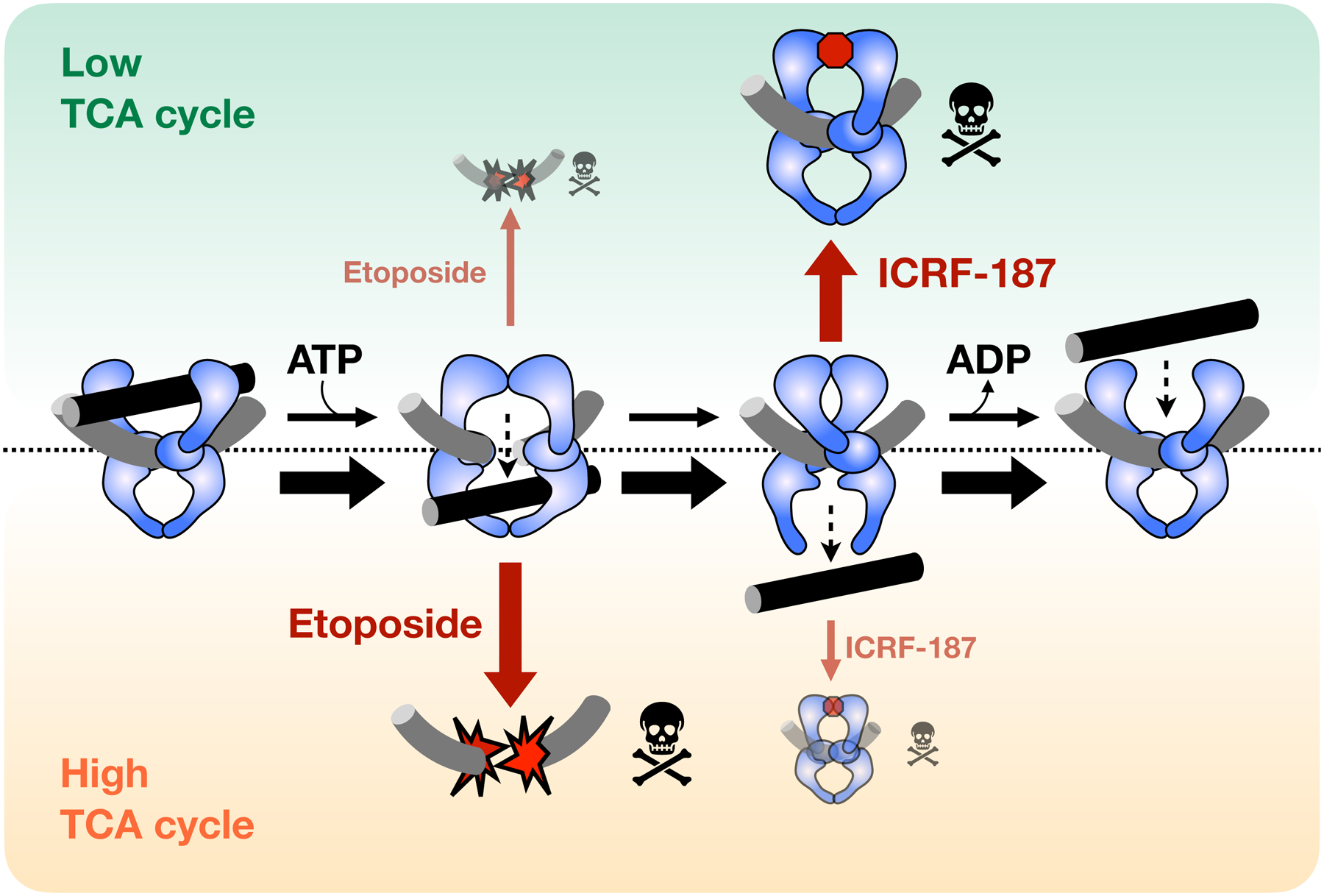
Topoisomerase II (topo II) is essential for disentangling newly replicated chromosomes. DNA unlinking involves the physical passage of one duplex through another and depends on the transient formation of double-stranded DNA breaks, a step exploited by frontline chemotherapeutics to kill cancer cells. Although anti-topo II drugs are efficacious, they also elicit cytotoxic side effects in normal cells; insights into how topo II is regulated in different cellular contexts is essential to improve their targeted use. Using chemical fractionation and mass spectrometry, we have discovered that topo II is subject to metabolic control through the TCA cycle. We show that TCA metabolites stimulate topo II activity in vitro and that levels of TCA flux modulate cellular sensitivity to anti-topo II drugs in vivo. Our work reveals an unanticipated connection between the control of DNA topology and cellular metabolism, a finding with ramifications for the clinical use of anti-topo II therapies.
Keywords: DNA topology; ICRF-187; TCA cycle; cancer; chemotherapy; dexrazoxane; etoposide; metabolism; topoisomerase.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Anand R, Memisoglu G, and Haber J (2017). Cas9-mediated gene editing in Saccharomyces cerevisiae. Protoc. Exch 1–6.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
