

Dietary suppression of MHC class II expression in intestinal epithelial cells enhances intestinal tumorigenesis
- PMID: 34529935
- PMCID: PMC8650761
- DOI: 10.1016/j.stem.2021.08.007
Dietary suppression of MHC class II expression in intestinal epithelial cells enhances intestinal tumorigenesis
Abstract
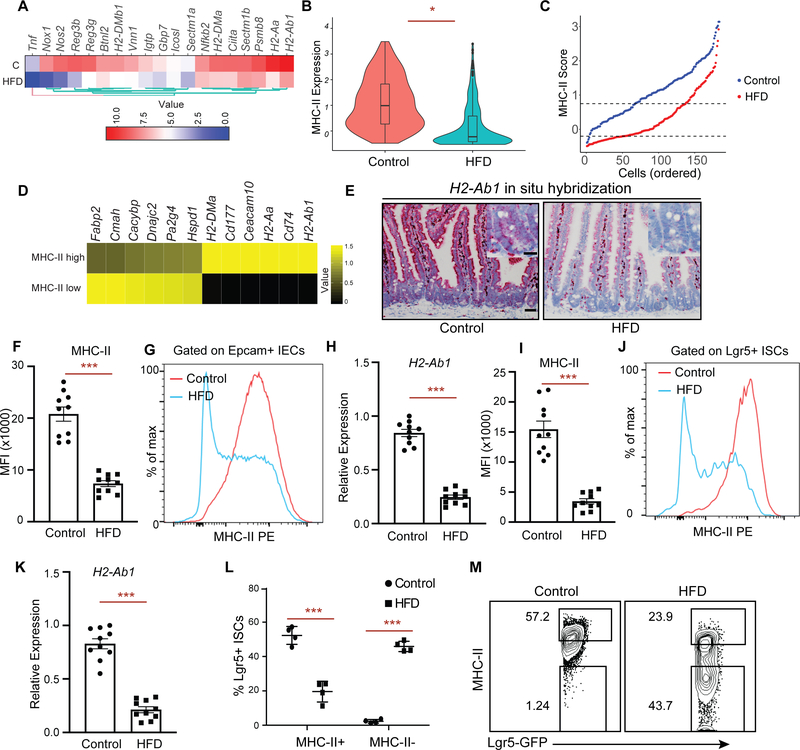
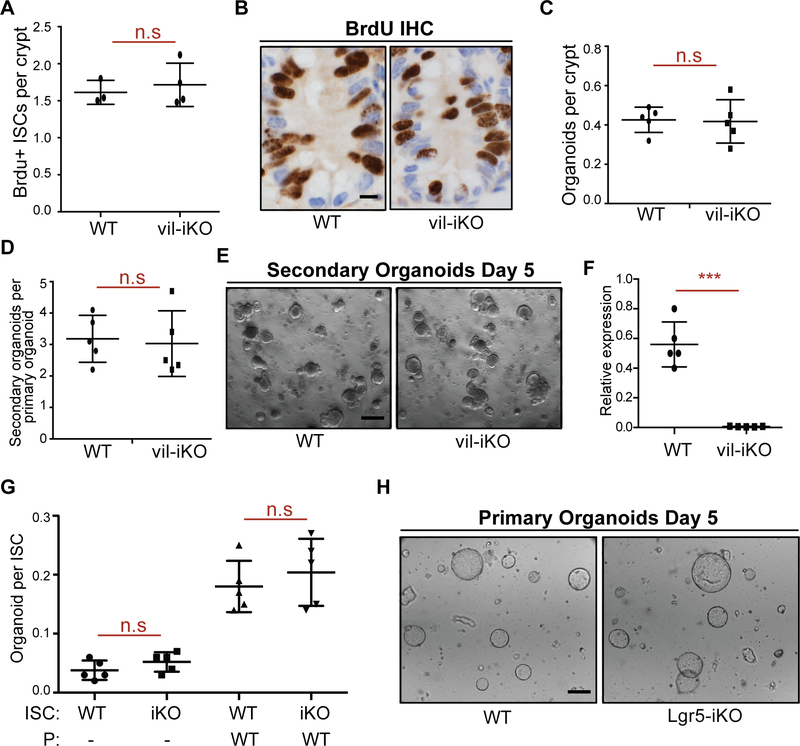
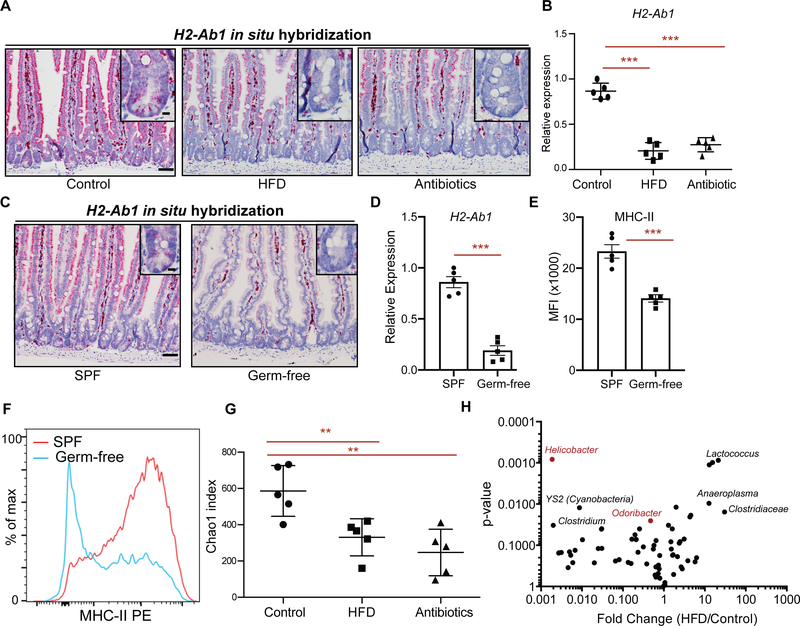
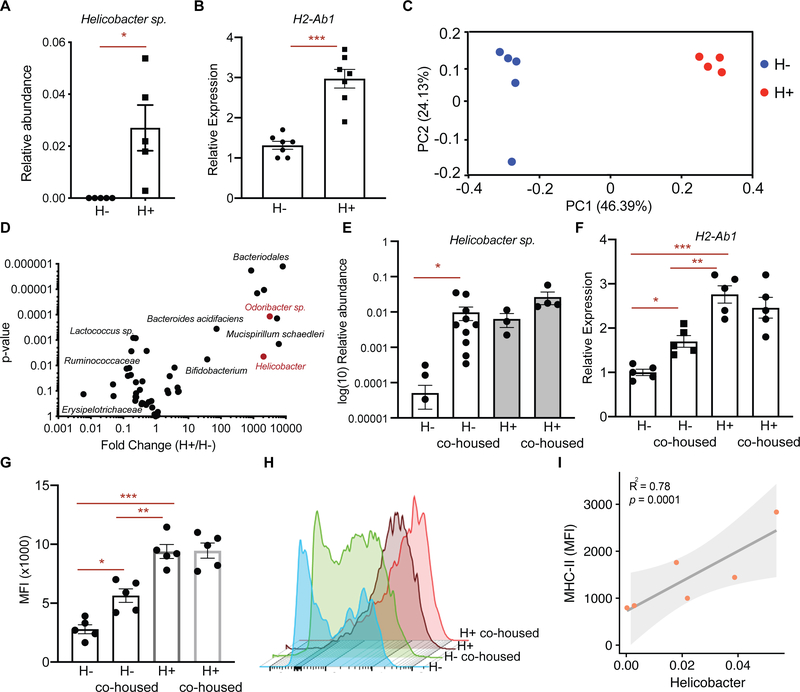
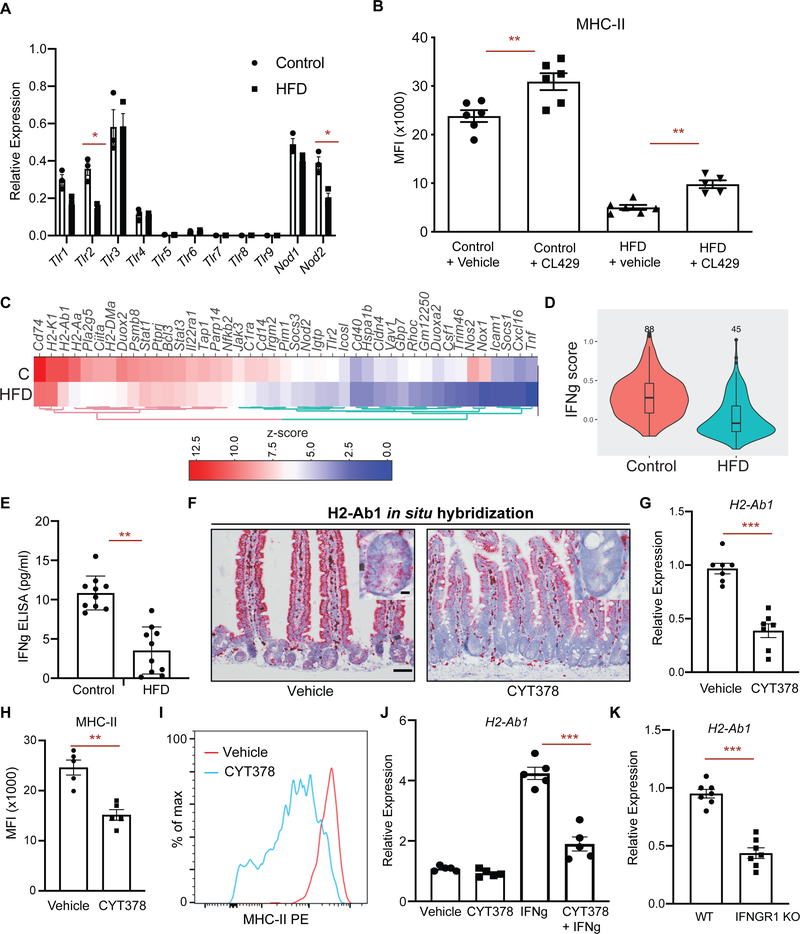
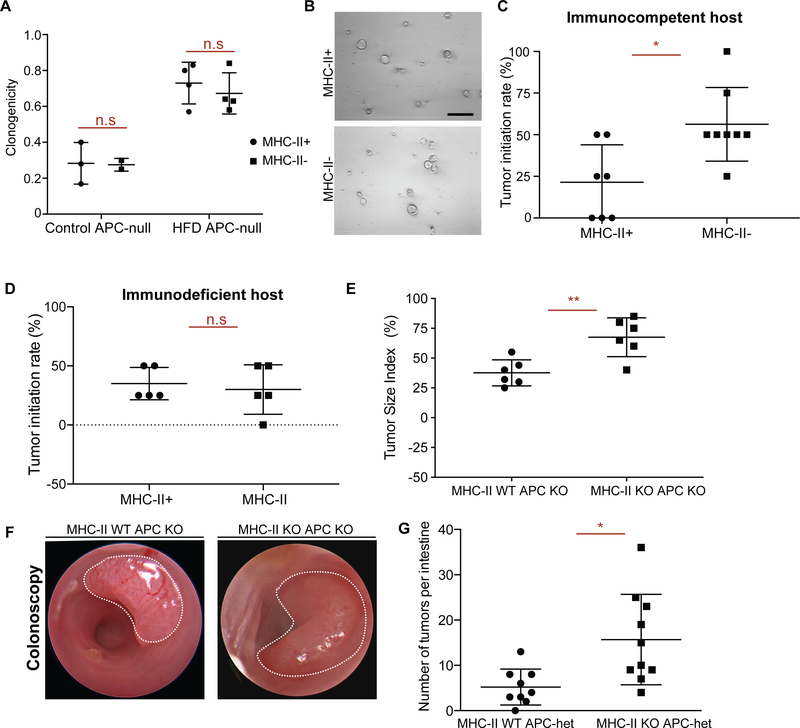
Little is known about how interactions of diet, intestinal stem cells (ISCs), and immune cells affect early-stage intestinal tumorigenesis. We show that a high-fat diet (HFD) reduces the expression of the major histocompatibility complex class II (MHC class II) genes in intestinal epithelial cells, including ISCs. This decline in epithelial MHC class II expression in a HFD correlates with reduced intestinal microbiome diversity. Microbial community transfer experiments suggest that epithelial MHC class II expression is regulated by intestinal flora. Mechanistically, pattern recognition receptor (PRR) and interferon-gamma (IFNγ) signaling regulates epithelial MHC class II expression. MHC class II-negative (MHC-II-) ISCs exhibit greater tumor-initiating capacity than their MHC class II-positive (MHC-II+) counterparts upon loss of the tumor suppressor Apc coupled with a HFD, suggesting a role for epithelial MHC class II-mediated immune surveillance in suppressing tumorigenesis. ISC-specific genetic ablation of MHC class II increases tumor burden cell autonomously. Thus, HFD perturbs a microbiome-stem cell-immune cell interaction that contributes to tumor initiation in the intestine.
Keywords: MHC-II; antigen presentation; cancer; diet; high-fat diet; intestinal stem cells; microbiome; obesity.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing financial interests. S.H.O. serves as an advisory board member for Cell Stem Cell. S.B. received research funding from Elstar Therapeutics and Revitope Oncology for research that is not related to this study.
Figures
Comment in
-
Microbiome effects on immune surveillance of intestinal tumours.Nat Rev Immunol. 2021 Nov;21(11):692-693. doi: 10.1038/s41577-021-00629-5. Nat Rev Immunol. 2021. PMID: 34535786 No abstract available.
-
High fat stems tumor immune surveillance.Cell Rep Med. 2021 Dec 21;2(12):100483. doi: 10.1016/j.xcrm.2021.100483. eCollection 2021 Dec 21. Cell Rep Med. 2021. PMID: 35028622 Free PMC article.
References
-
- Abreu MT (2010). Toll-like receptor signalling in the intestinal epithelium: how bacterial recognition shapes intestinal function. Nat Rev Immunol 10, 131–144. - PubMed
-
- Barker N, Ridgway RA, van Es JH, van de Wetering M, Begthel H, van den Born M, Danenberg E, Clarke AR, Sansom OJ, and Clevers H (2009). Crypt stem cells as the cells-of-origin of intestinal cancer. Nature 457, 608–611. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
