

Metabolic actions of kisspeptin signaling: Effects on body weight, energy expenditure, and feeding
- PMID: 34530008
- PMCID: PMC8884343
- DOI: 10.1016/j.pharmthera.2021.107974
Metabolic actions of kisspeptin signaling: Effects on body weight, energy expenditure, and feeding
Abstract
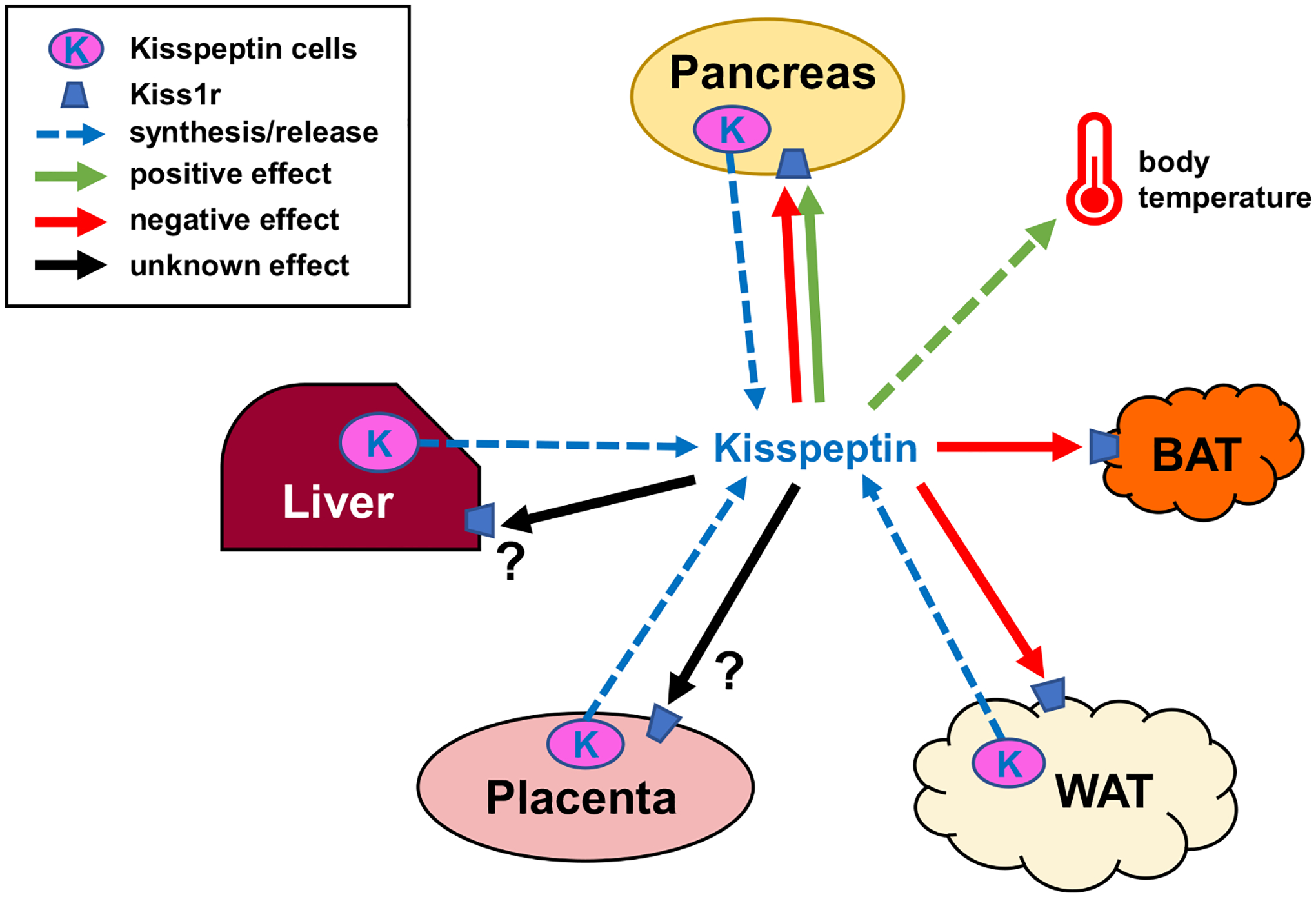
Kisspeptin (encoded by the Kiss1 gene) and its receptor, KISS1R (encoded by the Kiss1r gene), have well-established roles in stimulating reproduction via central actions on reproductive neural circuits, but recent evidence suggests that kisspeptin signaling also influences metabolism and energy balance. Indeed, both Kiss1 and Kiss1r are expressed in many metabolically-relevant peripheral tissues, including both white and brown adipose tissue, the liver, and the pancreas, suggesting possible actions on these tissues or involvement in their physiology. In addition, there may be central actions of kisspeptin signaling, or factors co-released from kisspeptin neurons, that modulate metabolic, feeding, or thermoregulatory processes. Accumulating data from animal models suggests that kisspeptin signaling regulates a wide variety of metabolic parameters, including body weight and energy expenditure, adiposity and adipose tissue function, food intake, glucose metabolism, respiratory rates, locomotor activity, and thermoregulation. Herein, the current evidence for the involvement of kisspeptin signaling in each of these physiological parameters is reviewed, gaps in knowledge identified, and future avenues of important research highlighted. Collectively, the discussed findings highlight emerging non-reproductive actions of kisspeptin signaling in metabolism and energy balance, in addition to previously documented roles in reproductive control, but also emphasize the need for more research to resolve current controversies and uncover underlying molecular and physiological mechanisms.
Keywords: Adipose; Body weight; Diabetes; Energy expenditure; Fat; Feeding; Food intake; GPR54; Insulin; KISS1; KISS1R; Kisspeptin; Metabolism; Obesity.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest
Both ADH and ASK declare no conflicts of interest.
Figures
Similar articles
-
Gonadal hormone-dependent vs. -independent effects of kisspeptin signaling in the control of body weight and metabolic homeostasis.Metabolism. 2019 Sep;98:84-94. doi: 10.1016/j.metabol.2019.06.007. Epub 2019 Jun 19. Metabolism. 2019. PMID: 31226351
-
Conditional knockout of kisspeptin signaling in brown adipose tissue increases metabolic rate and body temperature and lowers body weight.FASEB J. 2020 Jan;34(1):107-121. doi: 10.1096/fj.201901600R. Epub 2019 Nov 19. FASEB J. 2020. PMID: 31914628 Free PMC article.
-
Novel actions of kisspeptin signaling outside of GnRH-mediated fertility: a potential role in energy balance.Domest Anim Endocrinol. 2020 Oct;73:106467. doi: 10.1016/j.domaniend.2020.106467. Epub 2020 Mar 5. Domest Anim Endocrinol. 2020. PMID: 32278499 Review.
-
The role of kisspeptin neurons in reproduction and metabolism.J Endocrinol. 2018 Sep;238(3):R173-R183. doi: 10.1530/JOE-18-0108. J Endocrinol. 2018. PMID: 30042117 Review.
-
Impaired kisspeptin signaling decreases metabolism and promotes glucose intolerance and obesity.J Clin Invest. 2014 Jul;124(7):3075-9. doi: 10.1172/JCI71075. Epub 2014 Jun 17. J Clin Invest. 2014. PMID: 24937427 Free PMC article.
Cited by
-
Effect of Kisspeptin-Type Neuropeptide on Locomotor Behavior and Muscle Physiology in the Sea Cucumber Apostichopus japonicus.Animals (Basel). 2023 Feb 17;13(4):705. doi: 10.3390/ani13040705. Animals (Basel). 2023. PMID: 36830492 Free PMC article.
-
Comprehensive chemoanatomical mapping, and the gonadal regulation, of seven kisspeptin neuronal populations in the mouse brain.J Neuroendocrinol. 2025 May;37(5):e70019. doi: 10.1111/jne.70019. Epub 2025 Mar 18. J Neuroendocrinol. 2025. PMID: 40102056 Free PMC article.
-
International society of sports nutrition position stand: nutritional concerns of the female athlete.J Int Soc Sports Nutr. 2023 Dec;20(1):2204066. doi: 10.1080/15502783.2023.2204066. J Int Soc Sports Nutr. 2023. PMID: 37221858 Free PMC article. Review.
-
Targeted inhibition of kisspeptin neurons reverses hyperandrogenemia and abnormal hyperactive LH secretion in a preclinical mouse model of polycystic ovary syndrome.Hum Reprod. 2024 Sep 1;39(9):2089-2103. doi: 10.1093/humrep/deae153. Hum Reprod. 2024. PMID: 38978296 Free PMC article.
-
Airway-associated adipose tissue accumulation is increased in a kisspeptin receptor knockout mouse model.Clin Sci (Lond). 2023 Oct 11;137(19):1547-1562. doi: 10.1042/CS20230792. Clin Sci (Lond). 2023. PMID: 37732890 Free PMC article.
References
-
- Babwah AV (2015). Uterine and placental KISS1 regulate pregnancy: What we know and the challenges that lie ahead. REPRODUCTION 150(4), R121. - PubMed
-
- Backholer K, et al. (2010). Kisspeptin cells in the ewe brain respond to leptin and communicate with neuropeptide Y and proopiomelanocortin cells. Endocrinology 151 (5), 2233–2243. - PubMed
-
- Baron AD, et al. (1987). Role of hyperglucagonemia in maintenance of increased rates of hepatic glucose output in type II diabetics. Diabetes 36(3), 274–283. - PubMed
