

Activation loop phosphorylation of a non-RD receptor kinase initiates plant innate immune signaling
- PMID: 34531323
- PMCID: PMC8463890
- DOI: 10.1073/pnas.2108242118
Activation loop phosphorylation of a non-RD receptor kinase initiates plant innate immune signaling
Abstract
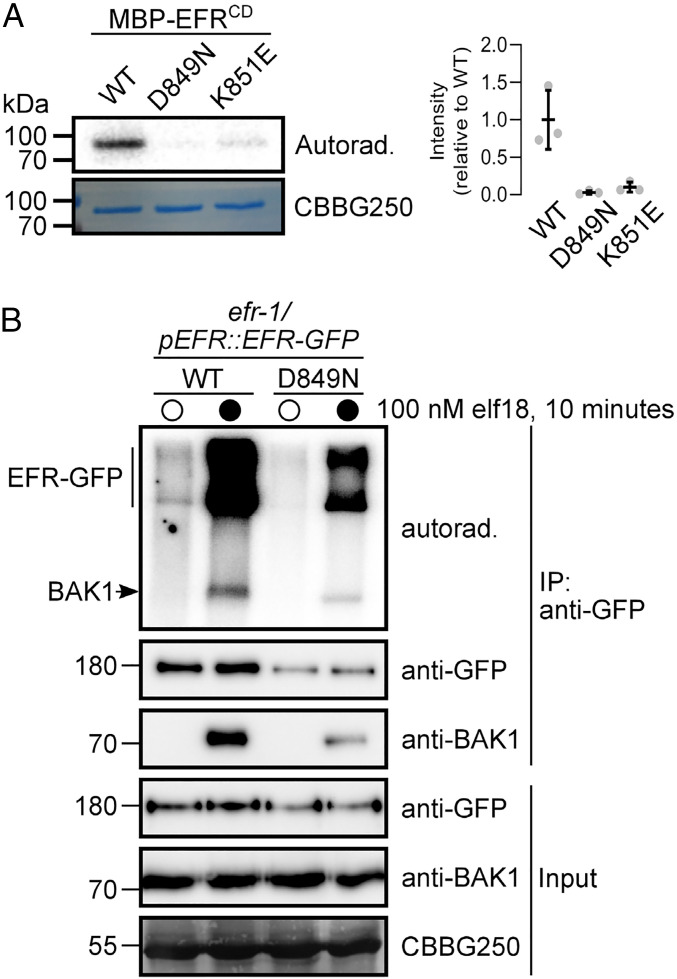
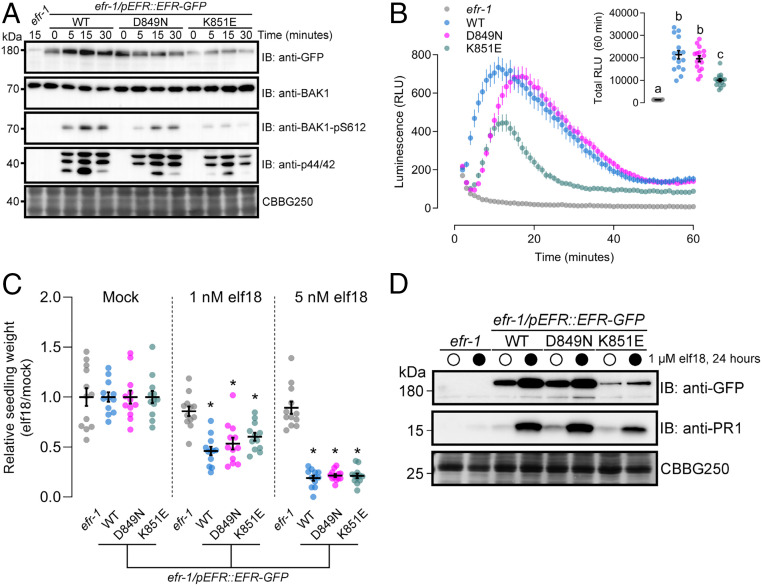
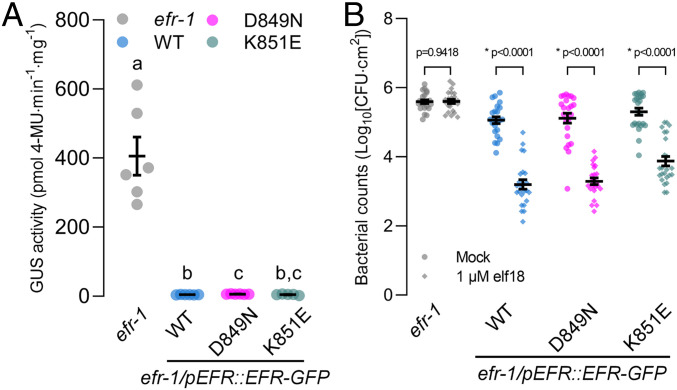
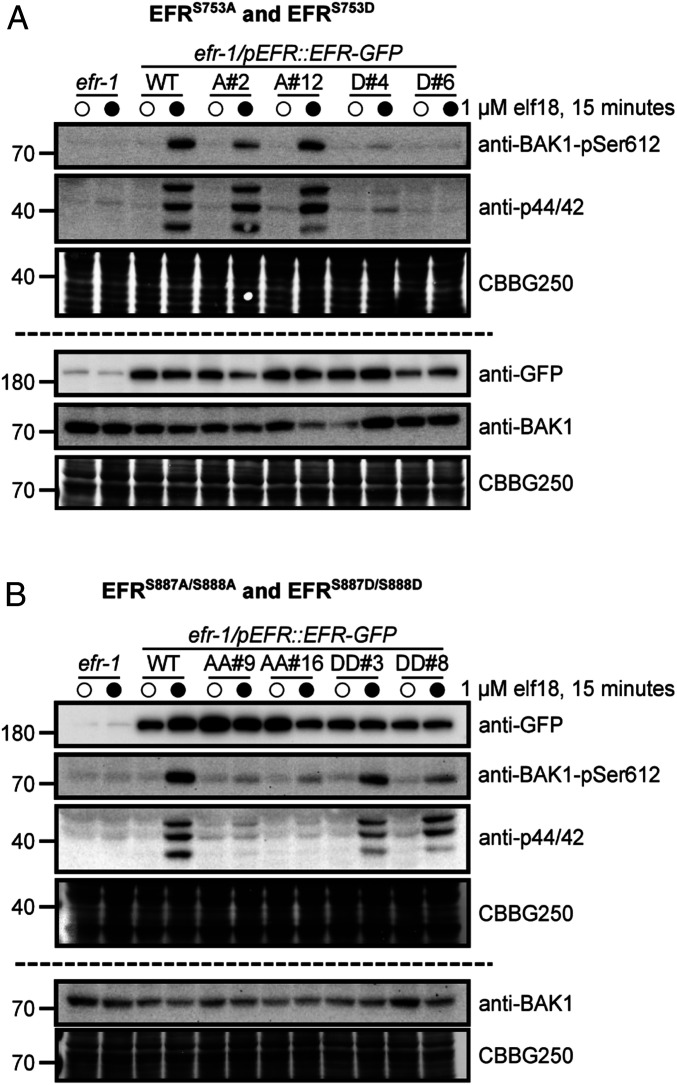
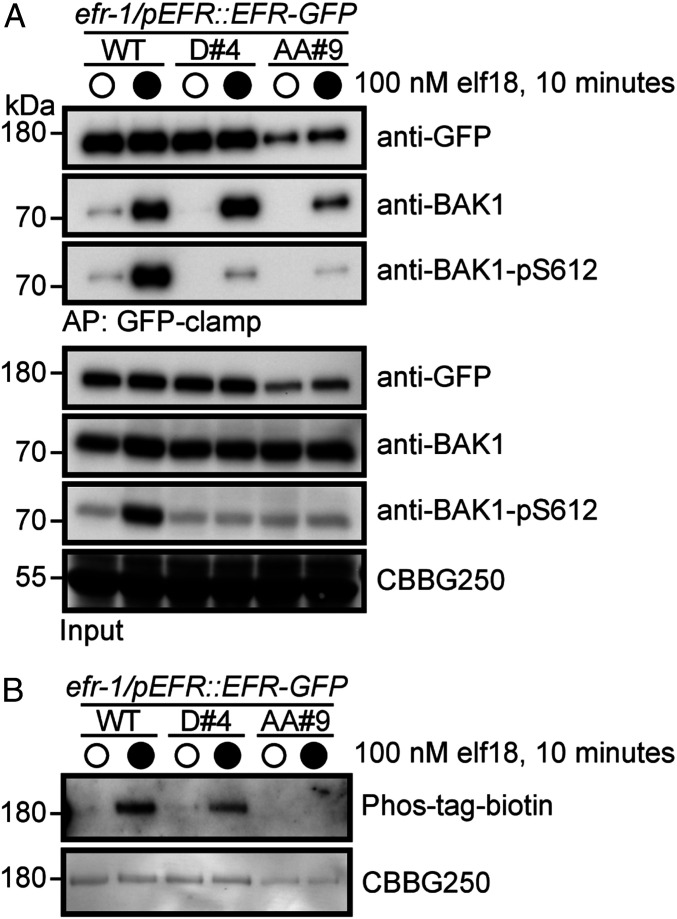
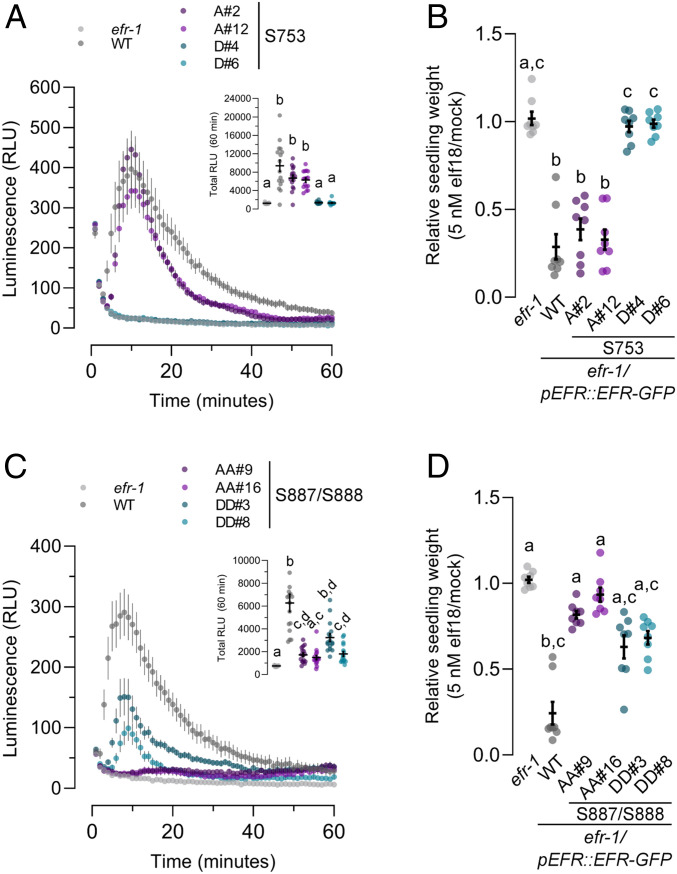
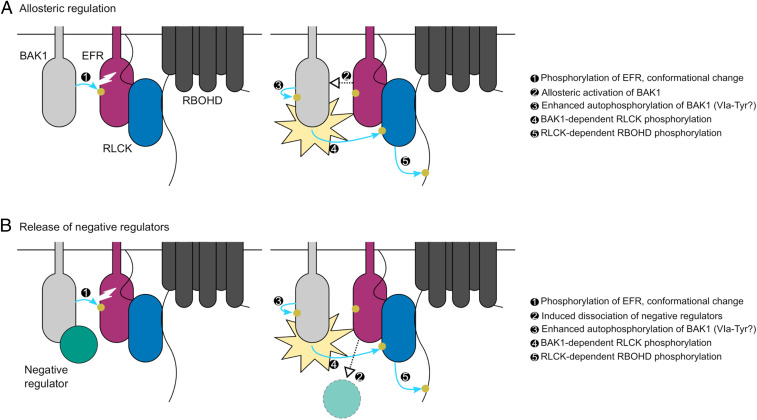
Receptor kinases (RKs) are fundamental for extracellular sensing and regulate development and stress responses across kingdoms. In plants, leucine-rich repeat receptor kinases (LRR-RKs) are primarily peptide receptors that regulate responses to myriad internal and external stimuli. Phosphorylation of LRR-RK cytoplasmic domains is among the earliest responses following ligand perception, and reciprocal transphosphorylation between a receptor and its coreceptor is thought to activate the receptor complex. Originally proposed based on characterization of the brassinosteroid receptor, the prevalence of complex activation via reciprocal transphosphorylation across the plant RK family has not been tested. Using the LRR-RK ELONGATION FACTOR TU RECEPTOR (EFR) as a model, we set out to understand the steps critical for activating RK complexes. While the EFR cytoplasmic domain is an active protein kinase in vitro and is phosphorylated in a ligand-dependent manner in vivo, catalytically deficient EFR variants are functional in antibacterial immunity. These results reveal a noncatalytic role for EFR in triggering immune signaling and indicate that reciprocal transphoshorylation is not a ubiquitous requirement for LRR-RK complex activation. Rather, our analysis of EFR along with a detailed survey of the literature suggests a distinction between LRR-RKs with RD- versus non-RD protein kinase domains. Based on newly identified phosphorylation sites that regulate the activation state of the EFR complex in vivo, we propose that LRR-RK complexes containing a non-RD protein kinase may be regulated by phosphorylation-dependent conformational changes of the ligand-binding receptor, which could initiate signaling either allosterically or through driving the dissociation of negative regulators of the complex.
Keywords: phosphorylation; receptor kinase; signaling.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Hohmann U., Lau K., Hothorn M., The structural basis of ligand perception and signal activation by receptor kinases. Annu. Rev. Plant Biol. 68, 109–137 (2017). - PubMed
-
- Dievart A., Gottin C., Périn C., Ranwez V., Chantret N., Origin and diversity of plant receptor-like kinases. Annu. Rev. Plant Biol. 71, 131–156 (2020). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
