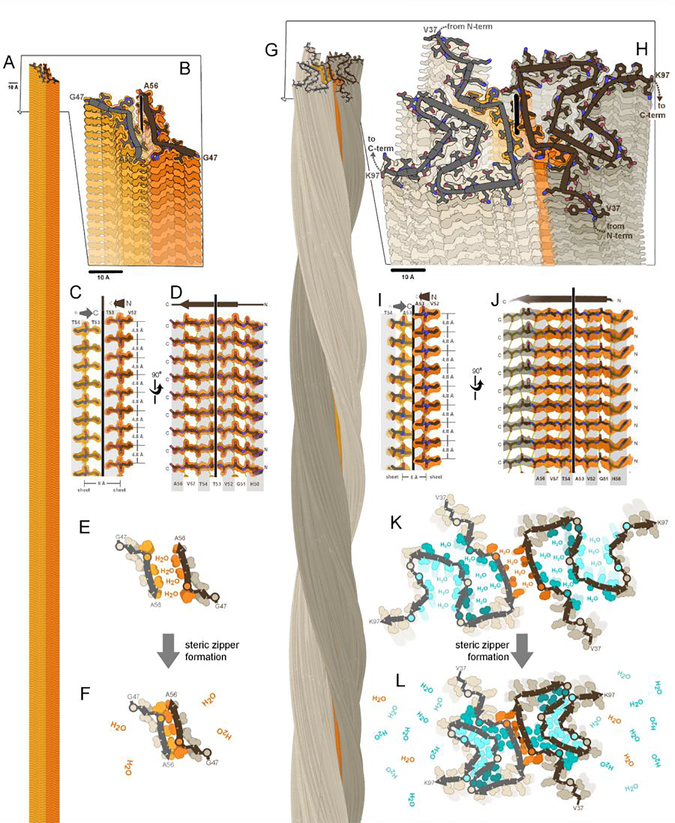
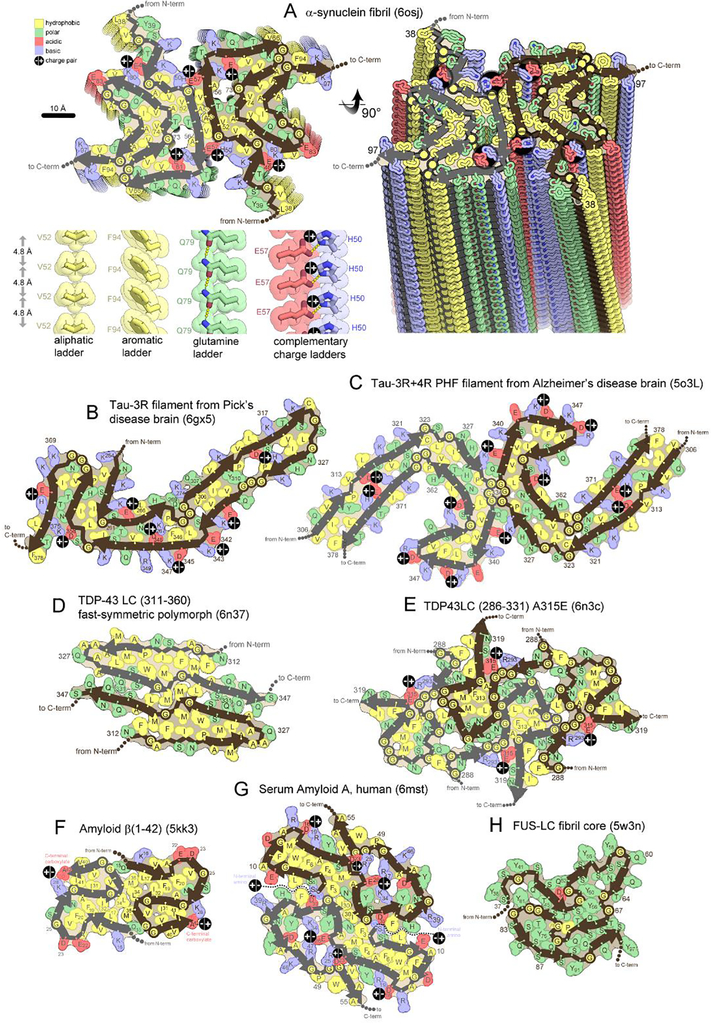
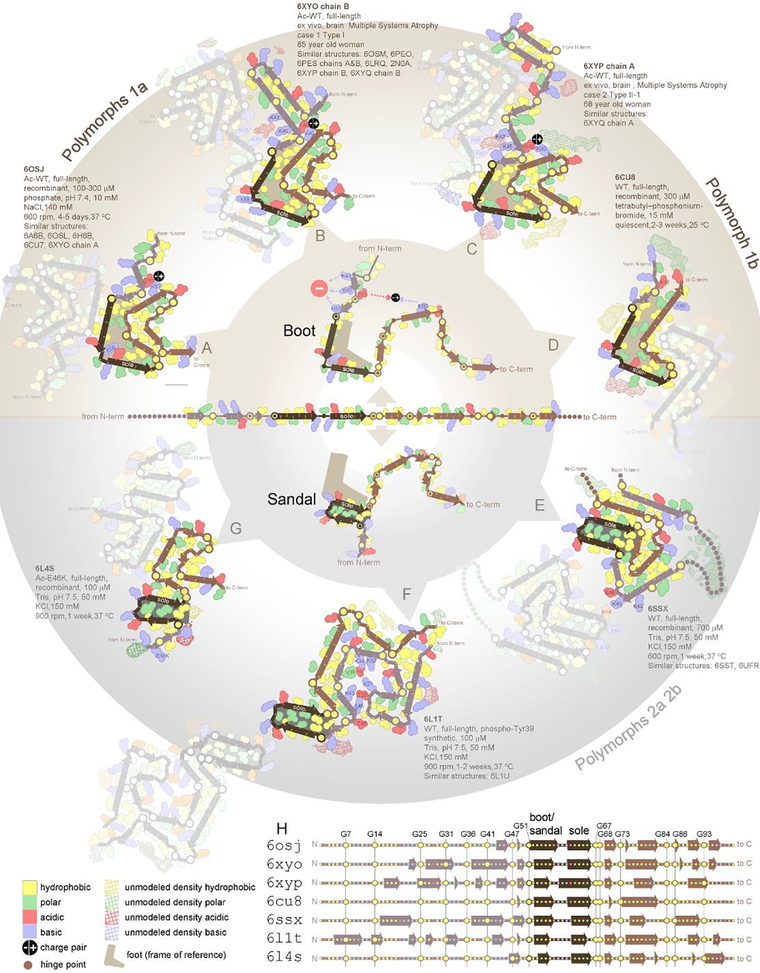
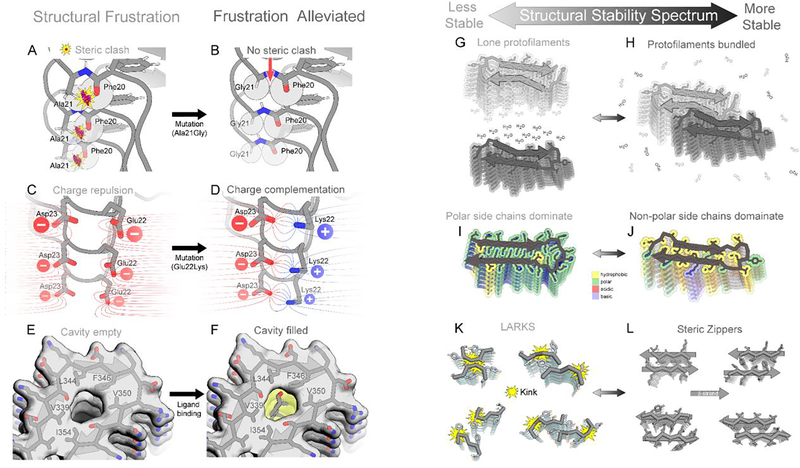
The expanding amyloid family: Structure, stability, function, and pathogenesis
- PMID: 34534463
- PMCID: PMC8772536
- DOI: 10.1016/j.cell.2021.08.013
The expanding amyloid family: Structure, stability, function, and pathogenesis
Abstract
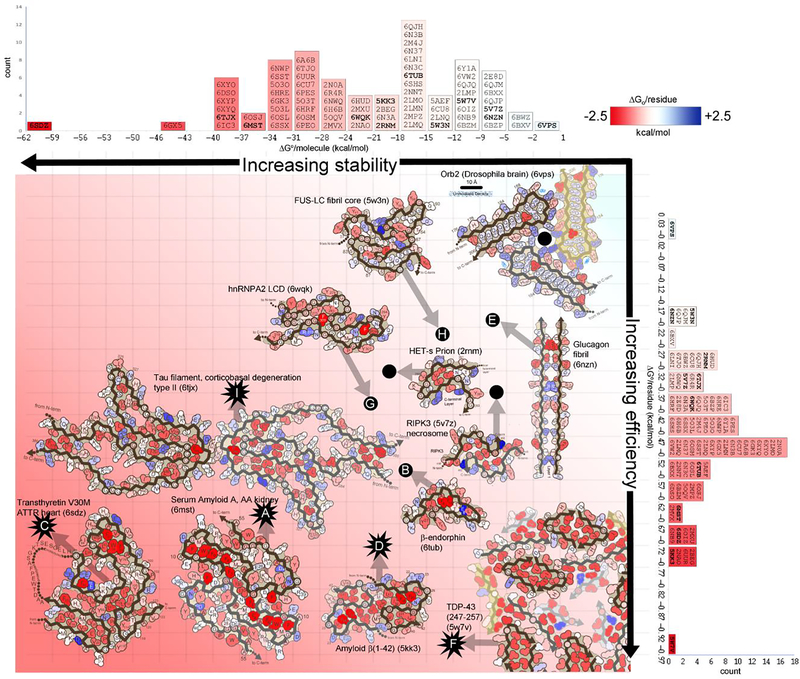
The hidden world of amyloid biology has suddenly snapped into atomic-level focus, revealing over 80 amyloid protein fibrils, both pathogenic and functional. Unlike globular proteins, amyloid proteins flatten and stack into unbranched fibrils. Stranger still, a single protein sequence can adopt wildly different two-dimensional conformations, yielding distinct fibril polymorphs. Thus, an amyloid protein may define distinct diseases depending on its conformation. At the heart of this conformational variability lies structural frustrations. In functional amyloids, evolution tunes frustration levels to achieve either stability or sensitivity according to the fibril's biological function, accounting for the vast versatility of the amyloid fibril scaffold.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests D.S.E. is SAB Chair and an equity holder in ADRx, Inc. J.A.R. is a founder and equity shareholder of Medstruc Inc.
Figures