

An extended DNA-free intranuclear compartment organizes centrosome microtubules in malaria parasites
- PMID: 34535568
- PMCID: PMC8473725
- DOI: 10.26508/lsa.202101199
An extended DNA-free intranuclear compartment organizes centrosome microtubules in malaria parasites
Abstract
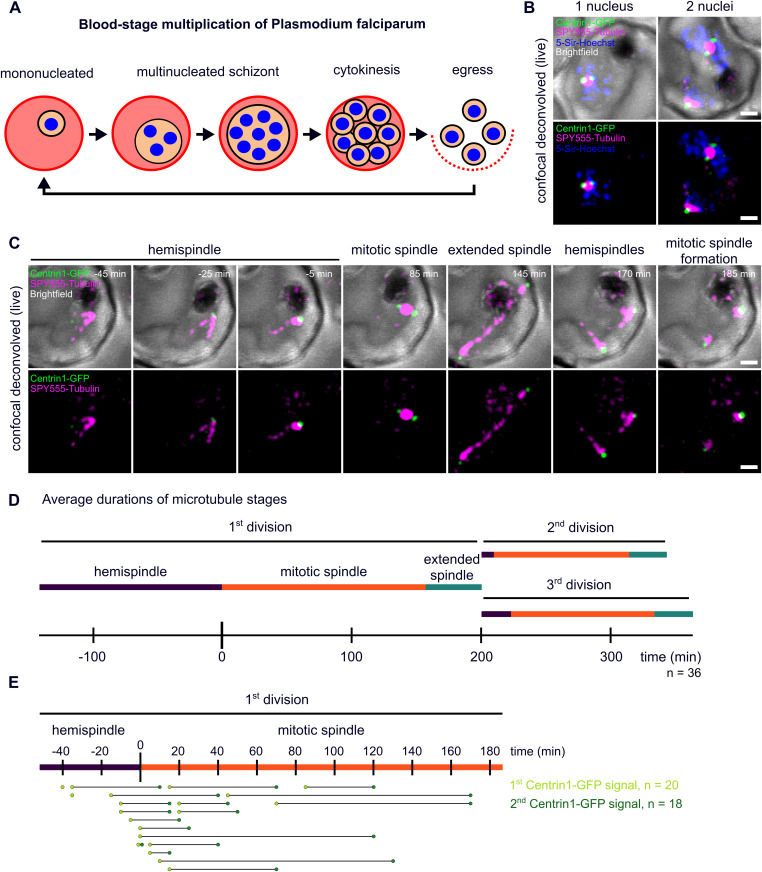
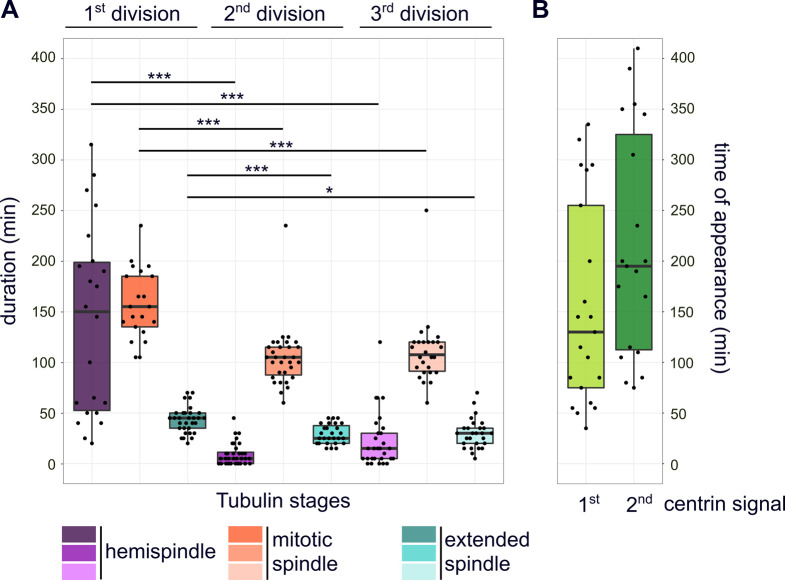
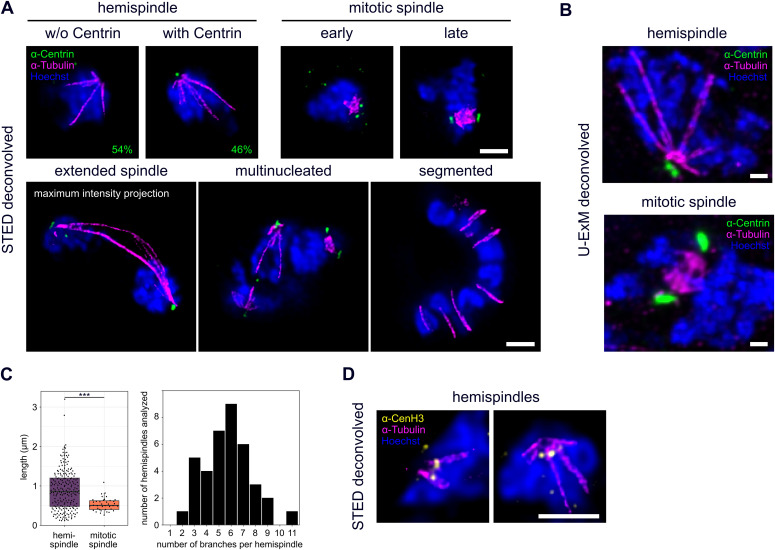
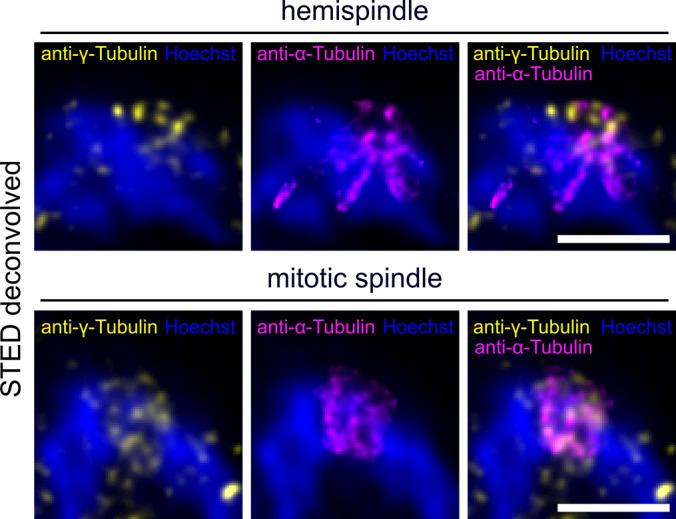
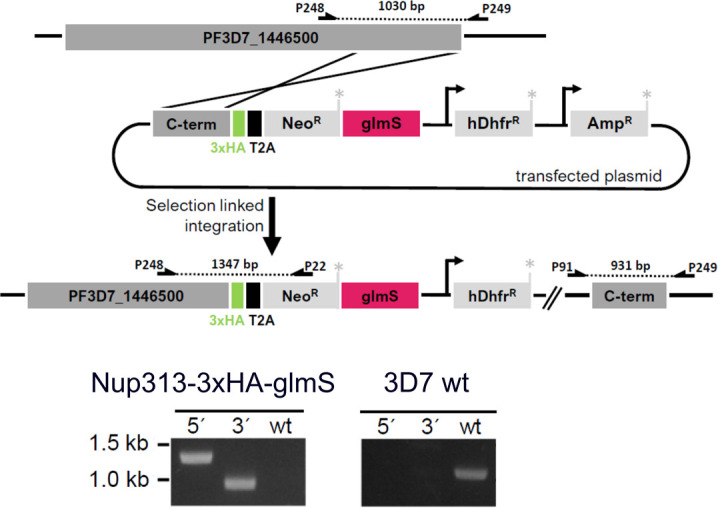
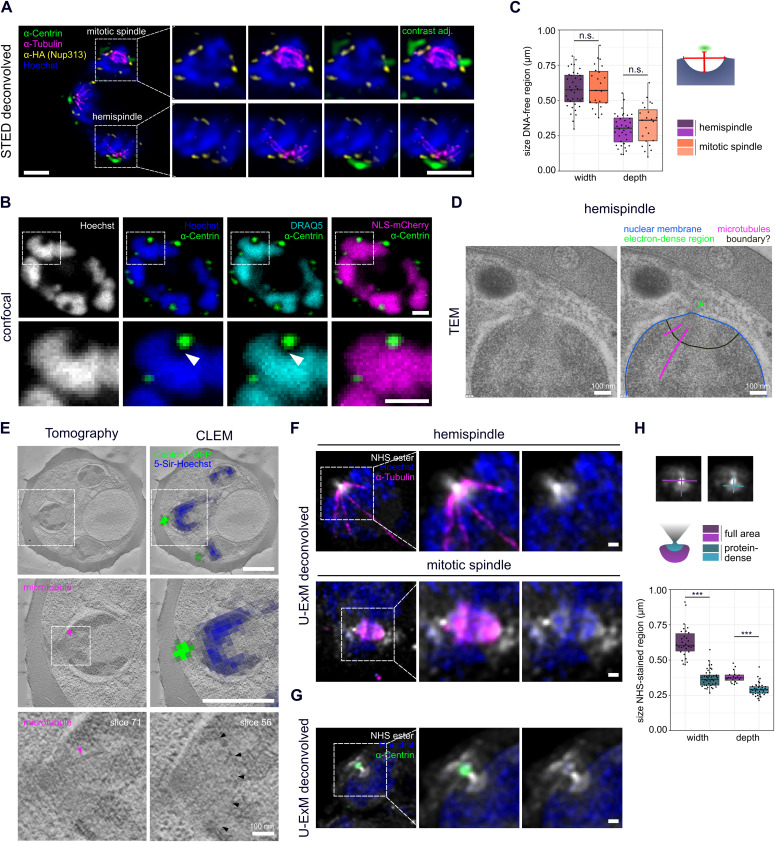
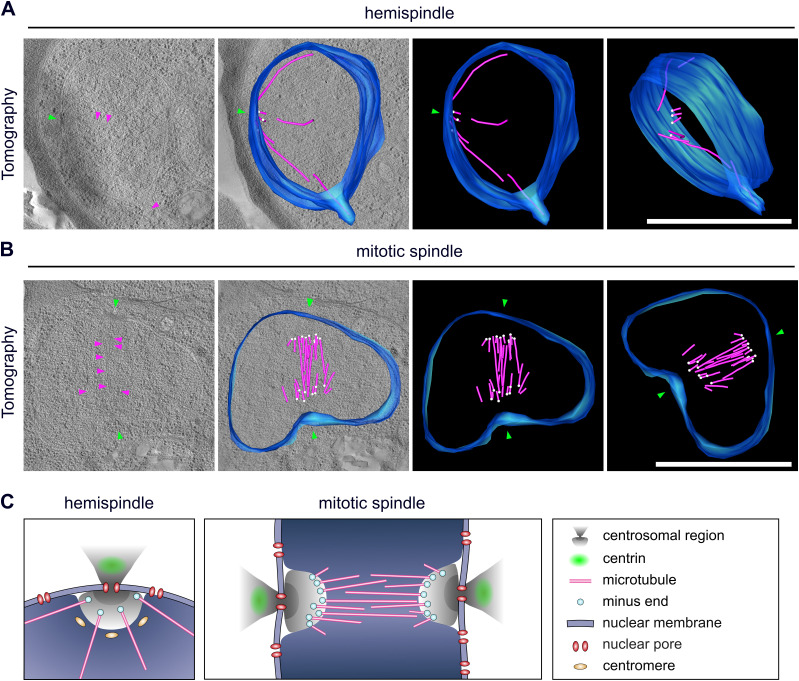
Proliferation of Plasmodium falciparum in red blood cells is the cause of malaria and is underpinned by an unconventional cell division mode, called schizogony. Contrary to model organisms, P. falciparum replicates by multiple rounds of nuclear divisions that are not interrupted by cytokinesis. Organization and dynamics of critical nuclear division factors remain poorly understood. Centriolar plaques, the centrosomes of P. falciparum, serve as microtubule organizing centers and have an acentriolar, amorphous structure. The small size of parasite nuclei has precluded detailed analysis of intranuclear microtubule organization by classical fluorescence microscopy. We apply recently developed super-resolution and time-lapse imaging protocols to describe microtubule reconfiguration during schizogony. Analysis of centrin, nuclear pore, and microtubule positioning reveals two distinct compartments of the centriolar plaque. Whereas centrin is extranuclear, we confirm by correlative light and electron tomography that microtubules are nucleated in a previously unknown and extended intranuclear compartment, which is devoid of chromatin but protein-dense. This study generates a working model for an unconventional centrosome and enables a better understanding about the diversity of eukaryotic cell division.
© 2021 Simon et al.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



Comment in
-
Hand-in-hand advances in microscopy and Plasmodium nuclear biology.Trends Parasitol. 2022 Jun;38(6):421-423. doi: 10.1016/j.pt.2022.03.007. Epub 2022 Apr 18. Trends Parasitol. 2022. PMID: 35450787
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources