

Sequencing refractory regions in bird genomes are hotspots for accelerated protein evolution
- PMID: 34537008
- PMCID: PMC8449477
- DOI: 10.1186/s12862-021-01905-7
Sequencing refractory regions in bird genomes are hotspots for accelerated protein evolution
Abstract
Background: Approximately 1000 protein encoding genes common for vertebrates are still unannotated in avian genomes. Are these genes evolutionary lost or are they not yet found for technical reasons? Using genome landscapes as a tool to visualize large-scale regional effects of genome evolution, we reexamined this question.
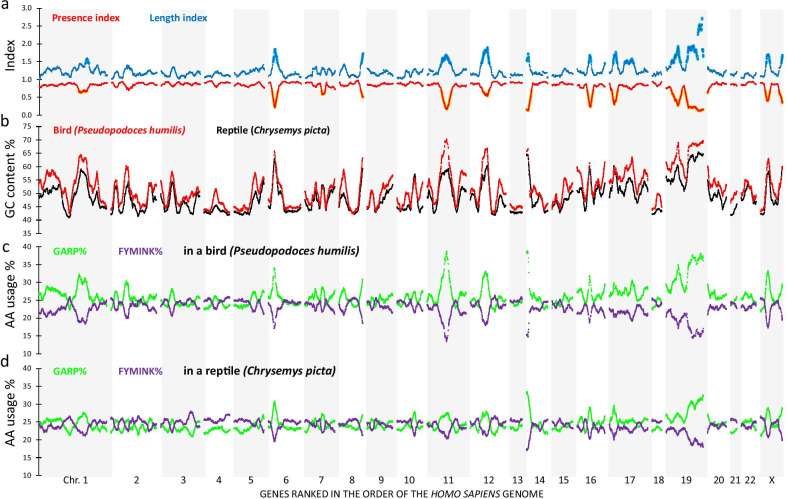
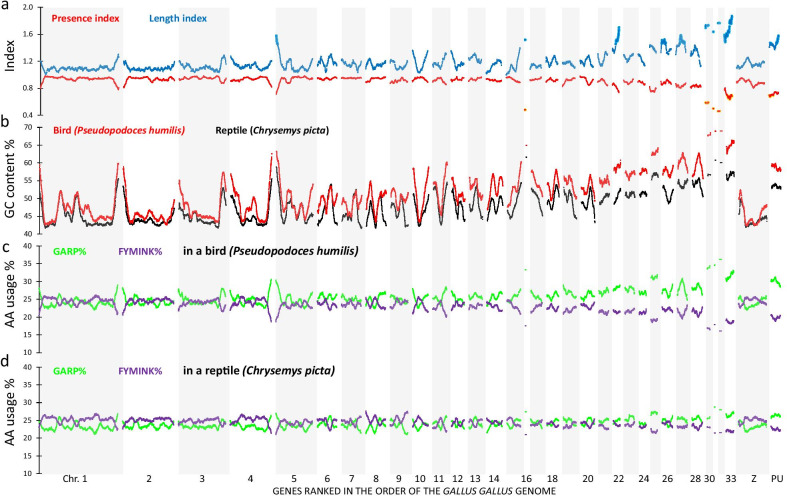
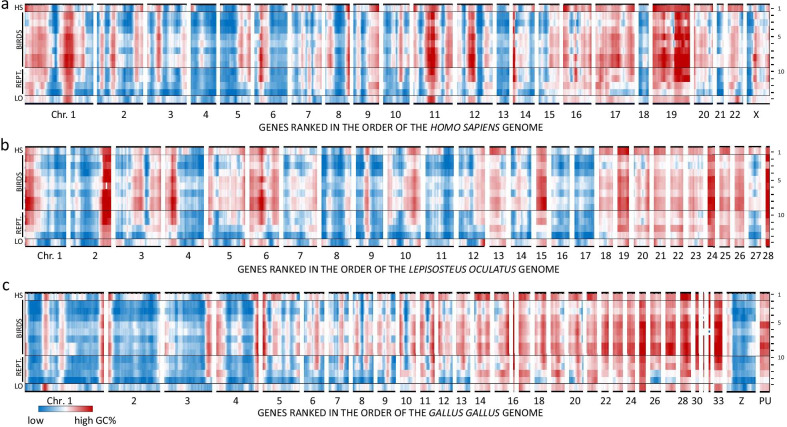
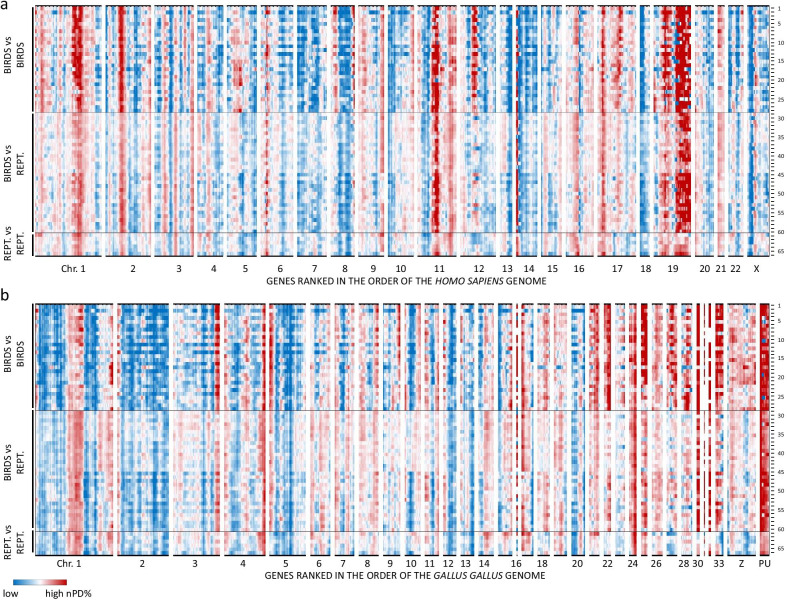
Results: On basis of gene annotation in non-avian vertebrate genomes, we established a list of 15,135 common vertebrate genes. Of these, 1026 were not found in any of eight examined bird genomes. Visualizing regional genome effects by our sliding window approach showed that the majority of these "missing" genes can be clustered to 14 regions of the human reference genome. In these clusters, an additional 1517 genes (often gene fragments) were underrepresented in bird genomes. The clusters of "missing" genes coincided with regions of very high GC content, particularly in avian genomes, making them "hidden" because of incomplete sequencing. Moreover, proteins encoded by genes in these sequencing refractory regions showed signs of accelerated protein evolution. As a proof of principle for this idea we experimentally characterized the mRNA and protein products of four "hidden" bird genes that are crucial for energy homeostasis in skeletal muscle: ALDOA, ENO3, PYGM and SLC2A4.
Conclusions: A least part of the "missing" genes in bird genomes can be attributed to an artifact caused by the difficulty to sequence regions with extreme GC% ("hidden" genes). Biologically, these "hidden" genes are of interest as they encode proteins that evolve more rapidly than the genome wide average. Finally we show that four of these "hidden" genes encode key proteins for energy metabolism in flight muscle.
Keywords: ALDOA; Accelerated; Avian genomes; ENO3; Evolution; GLUT4; Missing genes; PYGM; SLC2A4; Sequencing artifacts; Transcript landscapes.
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
-
- Carpenter KJ, Sutherland B. Eijkman’s contribution to the discovery of vitamins. J Nutr. 1995;125:155–163. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous