

Structures of synthetic nanobody-SARS-CoV-2 receptor-binding domain complexes reveal distinct sites of interaction
- PMID: 34537245
- PMCID: PMC8444450
- DOI: 10.1016/j.jbc.2021.101202
Structures of synthetic nanobody-SARS-CoV-2 receptor-binding domain complexes reveal distinct sites of interaction
Abstract
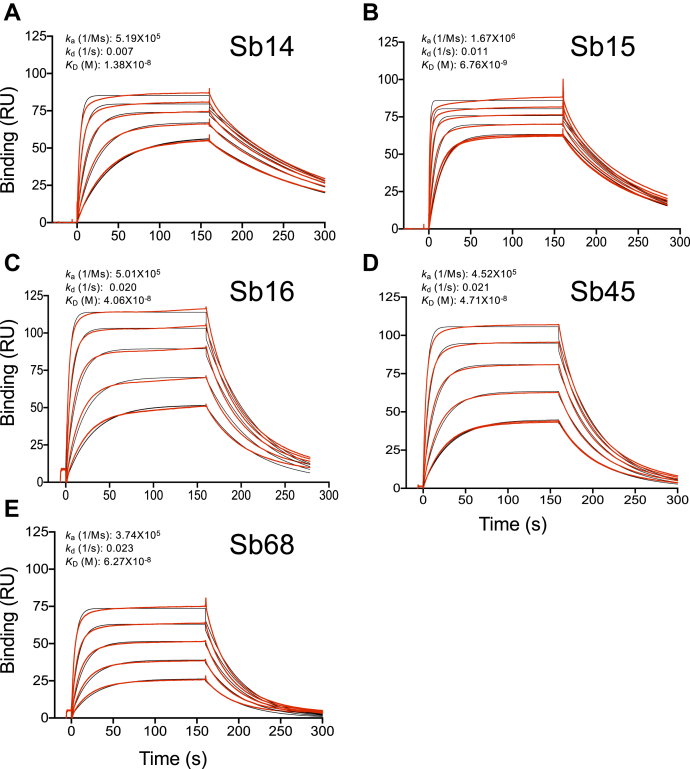
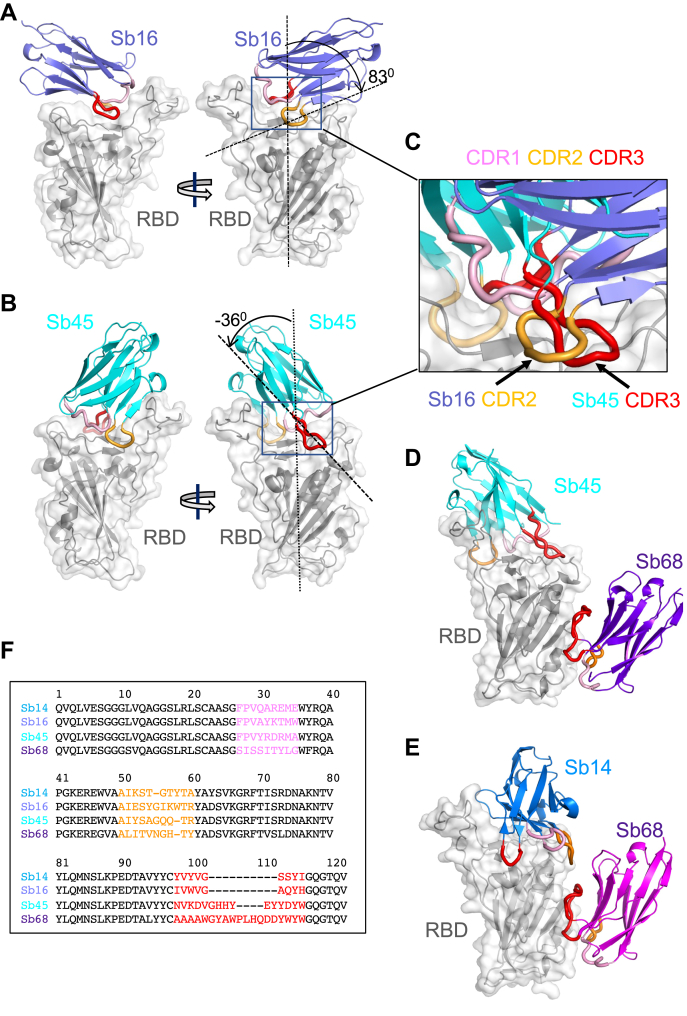
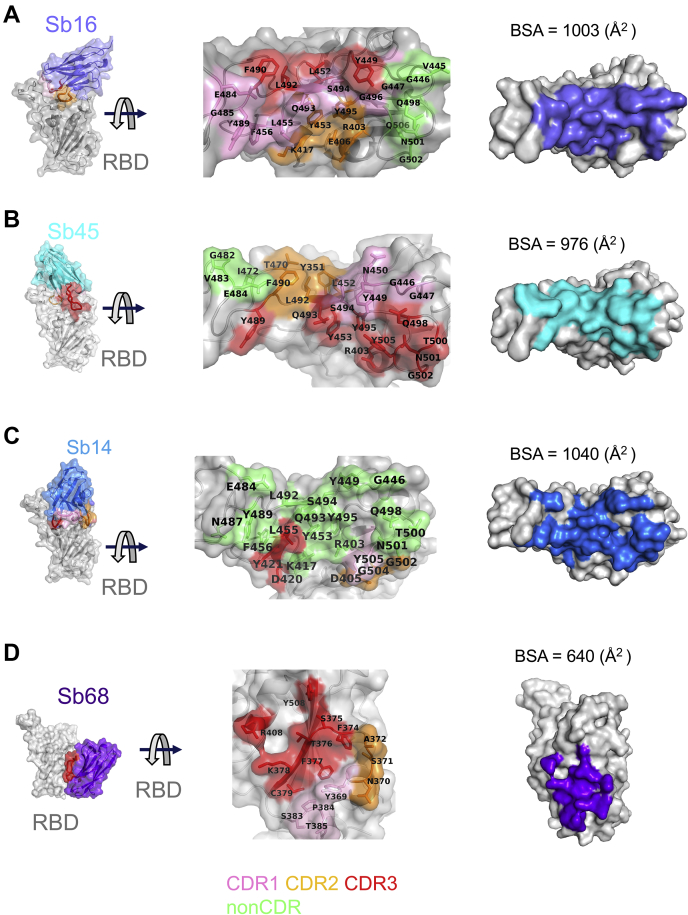
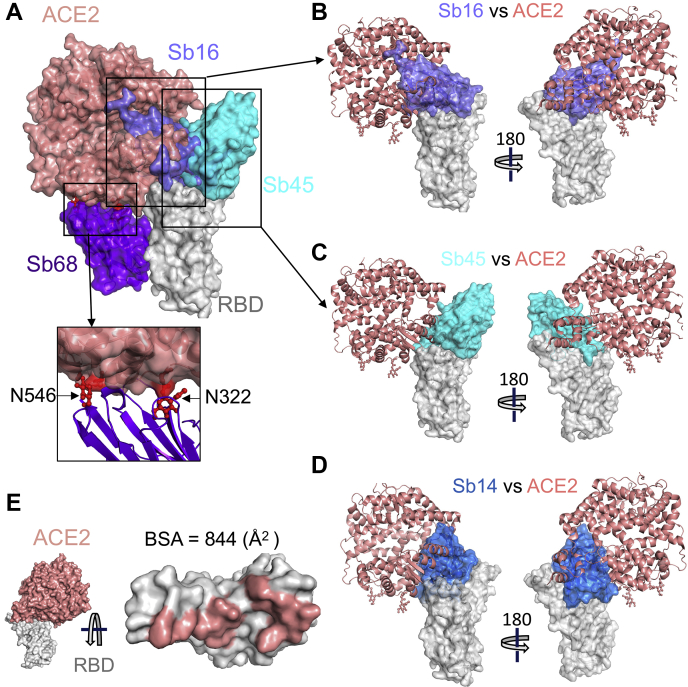
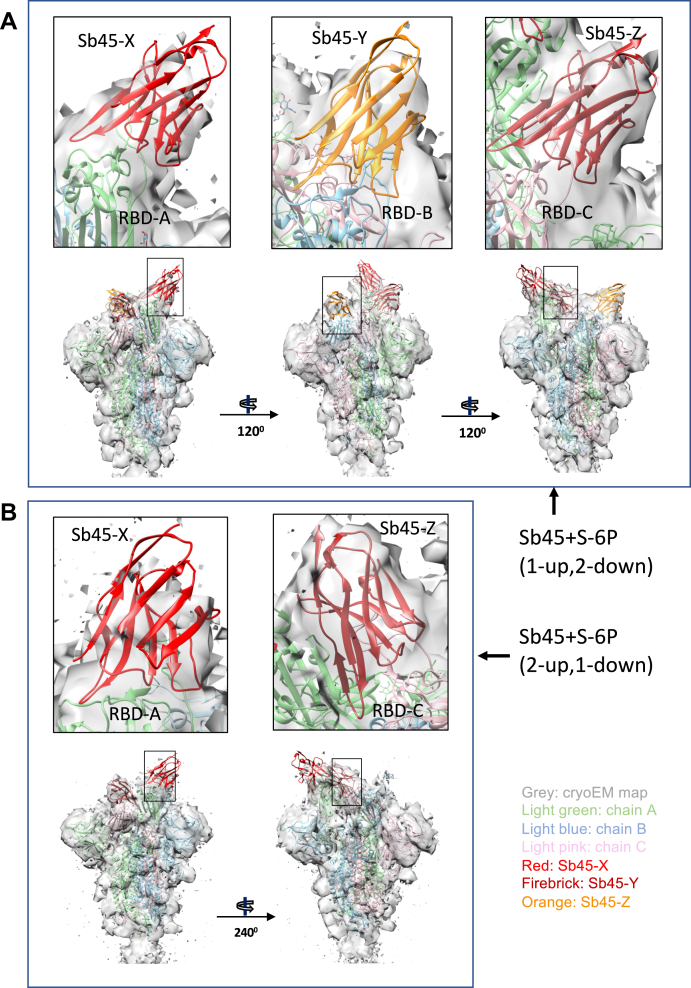
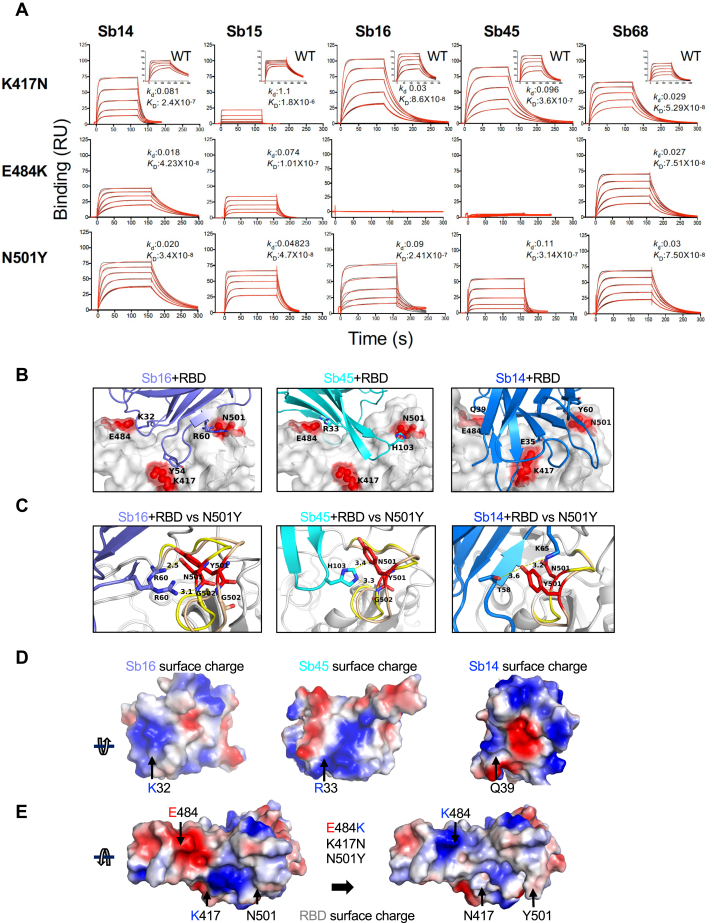
Combating the worldwide spread of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and the emergence of new variants demands understanding of the structural basis of the interaction of antibodies with the SARS-CoV-2 receptor-binding domain (RBD). Here, we report five X-ray crystal structures of sybodies (synthetic nanobodies) including those of binary and ternary complexes of Sb16-RBD, Sb45-RBD, Sb14-RBD-Sb68, and Sb45-RBD-Sb68, as well as unliganded Sb16. These structures reveal that Sb14, Sb16, and Sb45 bind the RBD at the angiotensin-converting enzyme 2 interface and that the Sb16 interaction is accompanied by a large conformational adjustment of complementarity-determining region 2. In contrast, Sb68 interacts at the periphery of the SARS-CoV-2 RBD-angiotensin-converting enzyme 2 interface. We also determined cryo-EM structures of Sb45 bound to the SARS-CoV-2 spike protein. Superposition of the X-ray structures of sybodies onto the trimeric spike protein cryo-EM map indicates that some sybodies may bind in both "up" and "down" configurations, but others may not. Differences in sybody recognition of several recently identified RBD variants are explained by these structures.
Keywords: SARS-CoV-2; cryo-EM; crystallography; single-domain antibody (sdAb, nanobody); surface plasmon resonance (SPR).
Published by Elsevier Inc.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
Update of
-
Synthetic nanobody-SARS-CoV-2 receptor-binding domain structures identify distinct epitopes.bioRxiv [Preprint]. 2021 Jan 27:2021.01.27.428466. doi: 10.1101/2021.01.27.428466. bioRxiv. 2021. Update in: J Biol Chem. 2021 Oct;297(4):101202. doi: 10.1016/j.jbc.2021.101202. PMID: 33532775 Free PMC article. Updated. Preprint.
-
Structures of synthetic nanobody-SARS-CoV-2-RBD complexes reveal distinct sites of interaction and recognition of variants.Res Sq [Preprint]. 2021 Jun 16:rs.3.rs-625642. doi: 10.21203/rs.3.rs-625642/v1. Res Sq. 2021. Update in: J Biol Chem. 2021 Oct;297(4):101202. doi: 10.1016/j.jbc.2021.101202. PMID: 34159326 Free PMC article. Updated. Preprint.
References
-
- Conti P., Caraffa A., Gallenga C.E., Kritas S.K., Frydas I., Younes A., Di Emidio P., Tete G., Pregliasco F., Ronconi G. The British variant of the new coronavirus-19 (Sars-Cov-2) should not create a vaccine problem. J. Biol. Regul. Homeost Agents. 2021;35:1–4. - PubMed
-
- Wibmer C.K., Ayres F., Hermanus T., Madzivhandila M., Kgagudi P., Oosthuysen B., Lambson B.E., de Oliveira T., Vermeulen M., van der Berg K., Rossouw T., Boswell M., Ueckermann V., Meiring S., von Gottberg A. SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. Nat. Med. 2021;27:622–625. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
