

EphA2 signaling within integrin adhesions regulates fibrillar adhesion elongation and fibronectin deposition
- PMID: 34537369
- PMCID: PMC8599653
- DOI: 10.1016/j.matbio.2021.09.001
EphA2 signaling within integrin adhesions regulates fibrillar adhesion elongation and fibronectin deposition
Abstract
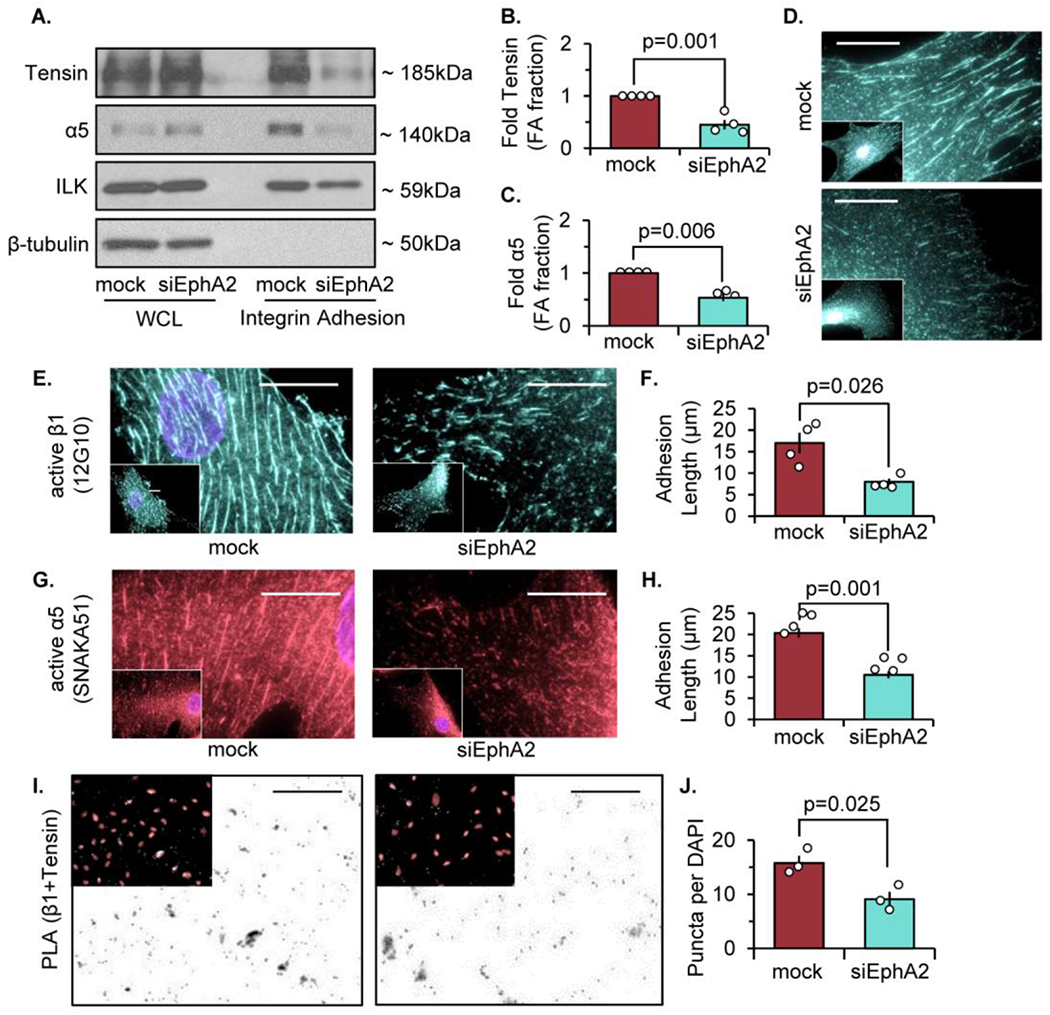
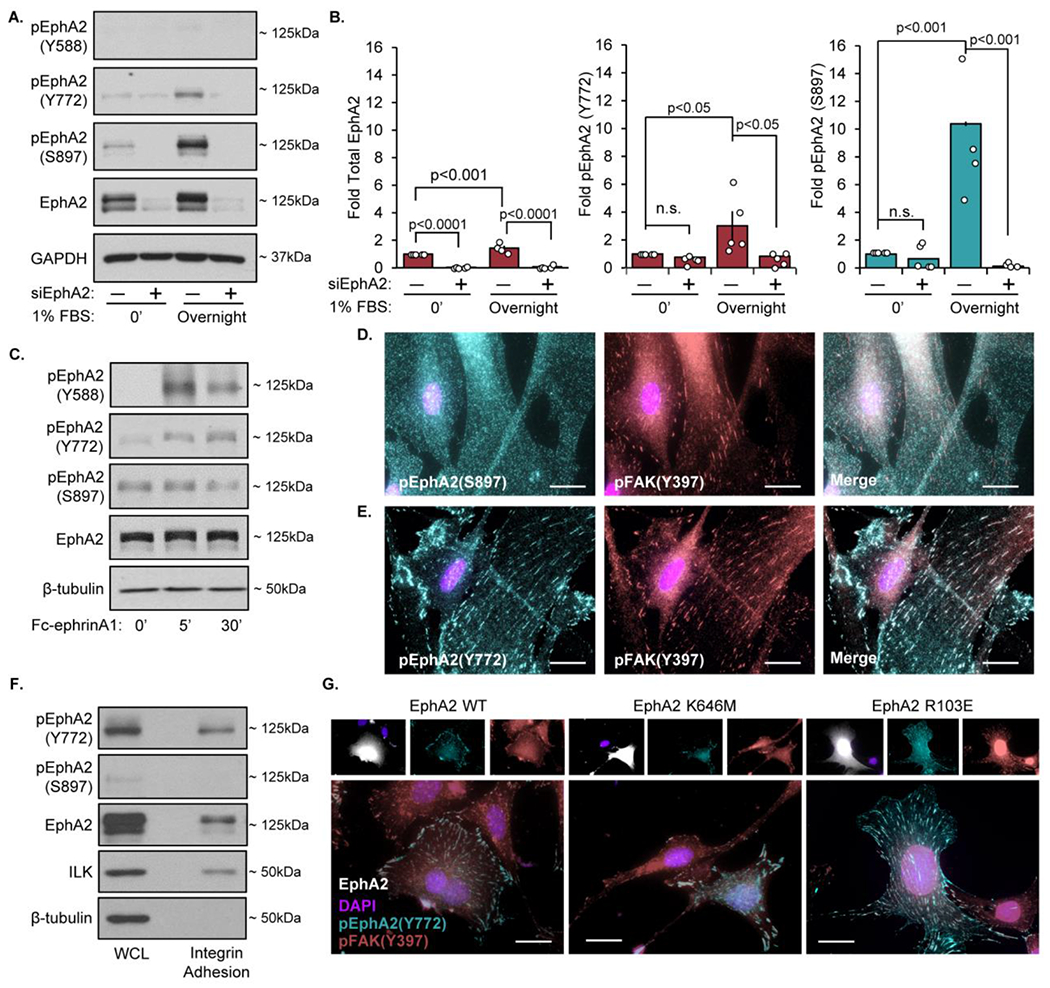
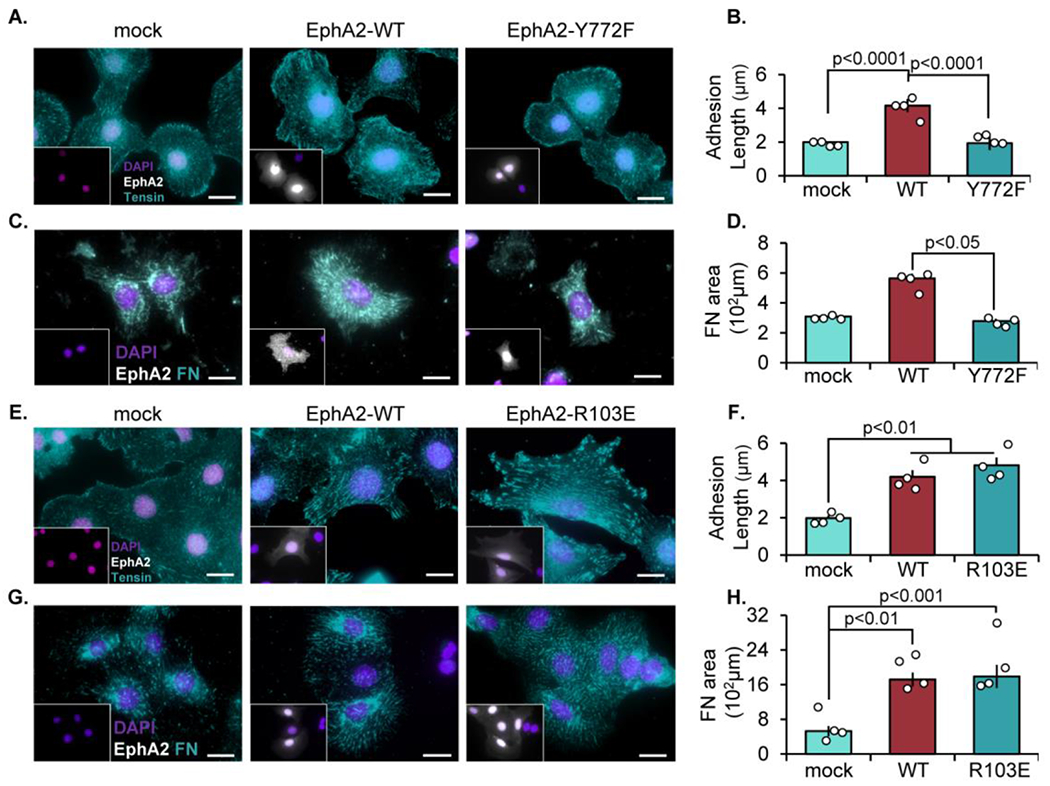
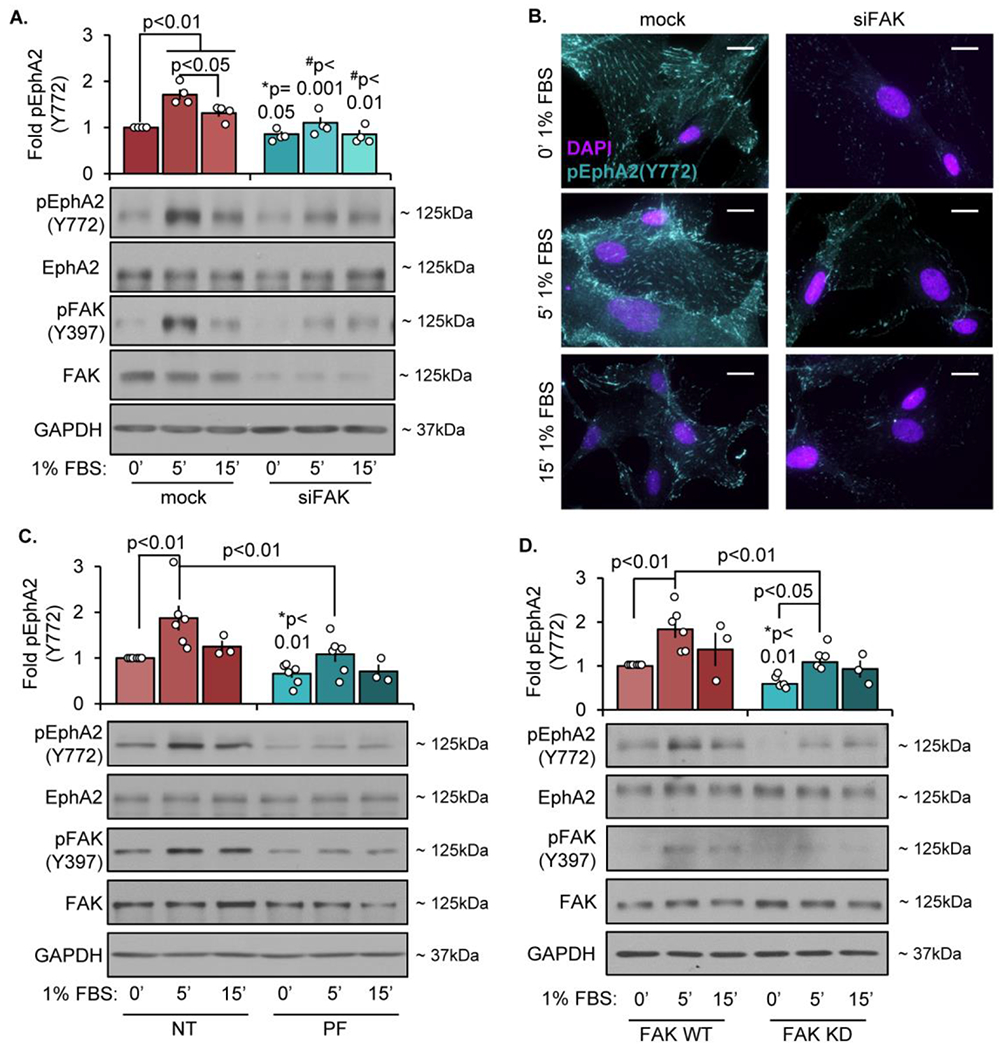
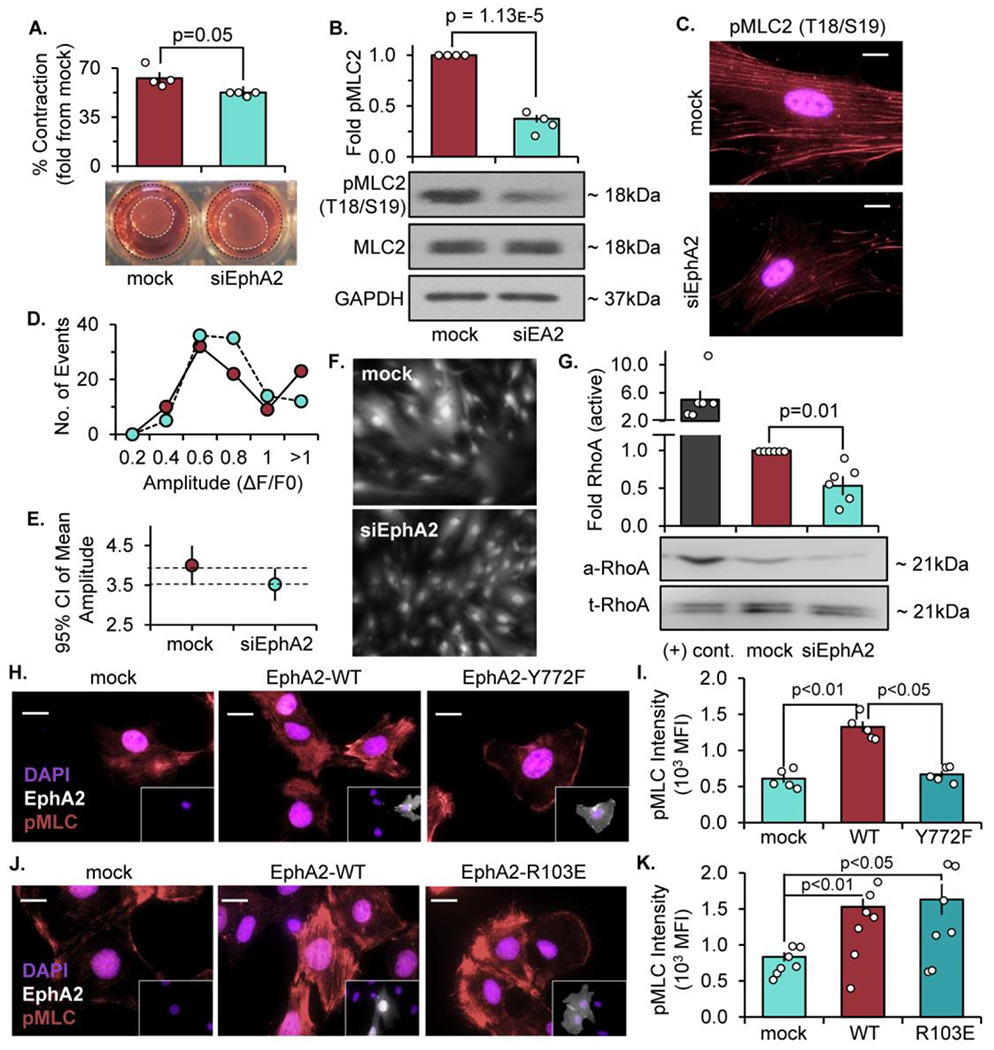
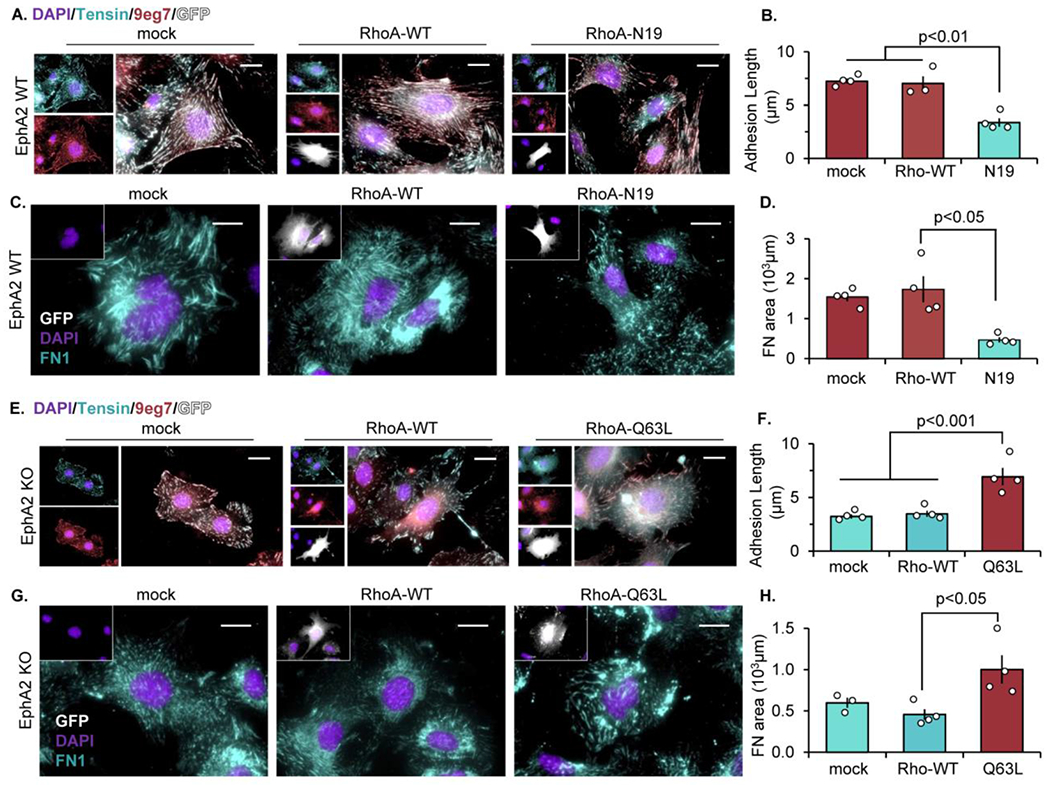
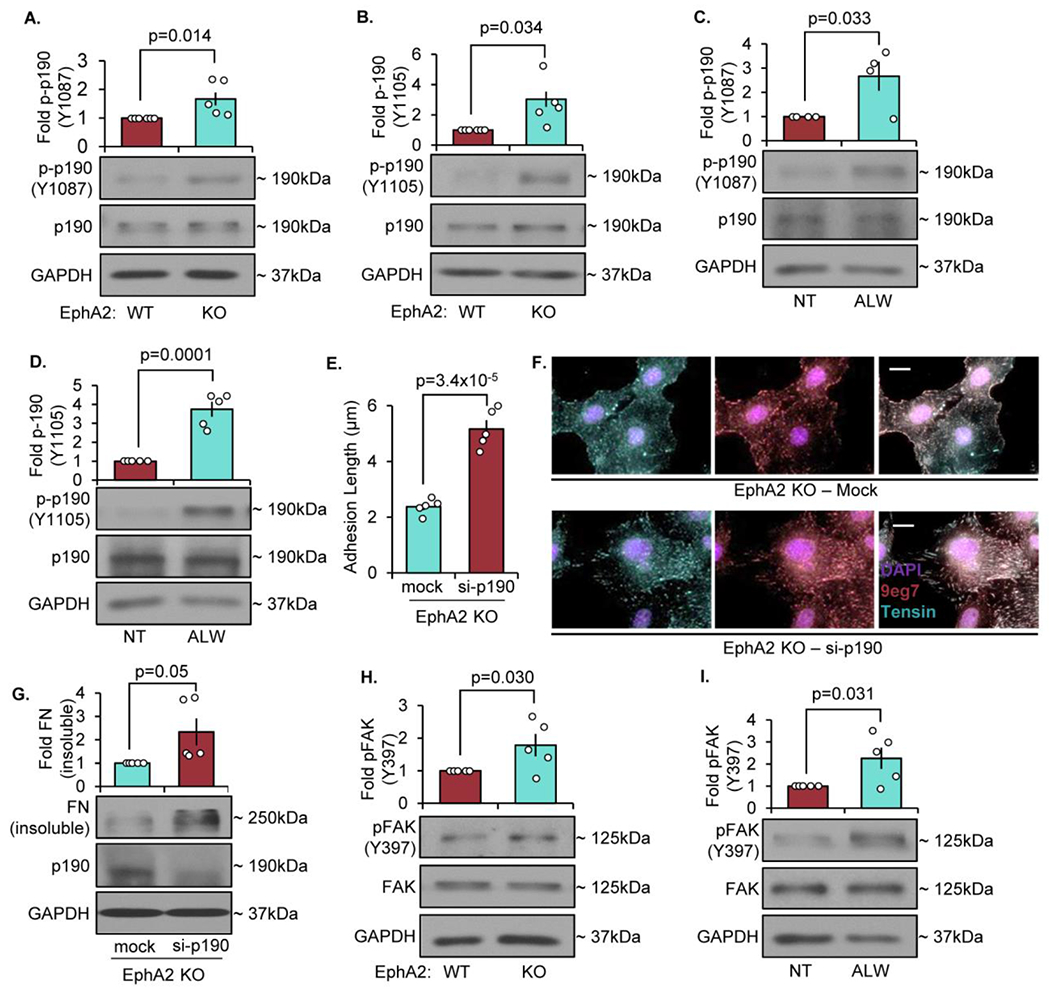
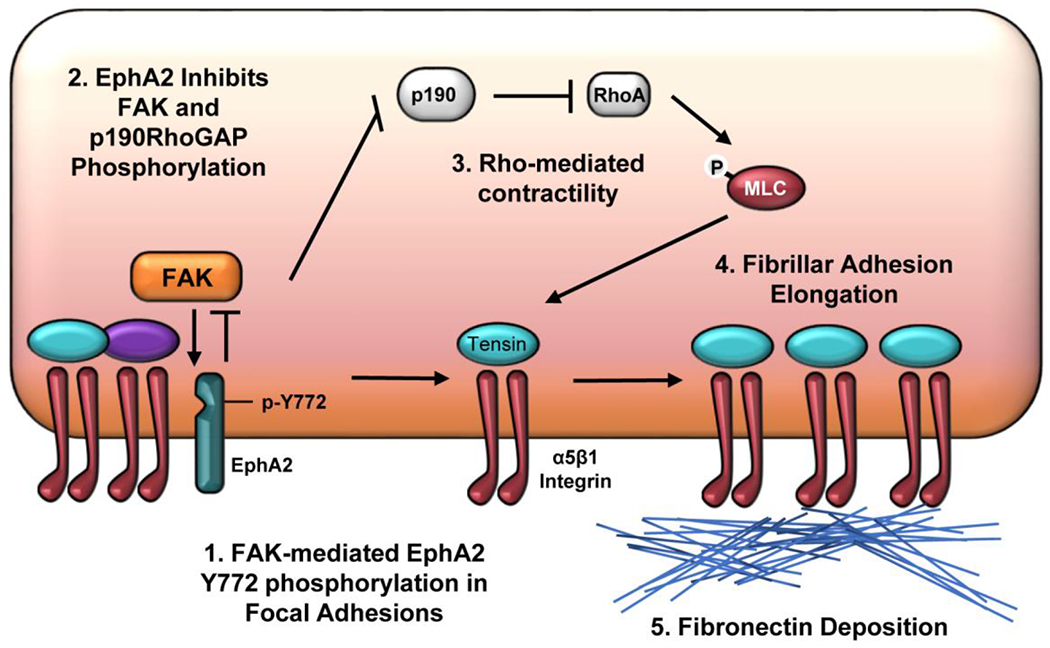
The multifunctional glycoprotein fibronectin influences several crucial cellular processes and contributes to multiple pathologies. While a link exists between fibronectin-associated pathologies and the receptor tyrosine kinase EphA2, the mechanism by which EphA2 promotes fibronectin matrix remodeling remains unknown. We previously demonstrated that EphA2 deletion reduces smooth muscle fibronectin deposition and blunts fibronectin deposition in atherosclerosis without influencing fibronectin expression. We now show that EphA2 expression is required for contractility-dependent elongation of tensin- and α5β1 integrin-rich fibrillar adhesions that drive fibronectin fibrillogenesis. Mechanistically, EphA2 localizes to integrin adhesions where focal adhesion kinase mediates ligand-independent Y772 phosphorylation, and mutation of this site significantly blunts fibrillar adhesion length. EphA2 deficiency decreases smooth muscle cell contractility by enhancing p190RhoGAP activation and reducing RhoA activity, whereas stimulating RhoA signaling in EphA2 deficient cells rescues fibrillar adhesion elongation. Together, these data identify EphA2 as a novel regulator of fibrillar adhesion elongation and provide the first data identifying a role for EphA2 signaling in integrin adhesions.
Keywords: EphA2 receptor tyrosine kinase; Fibrillar adhesions; Fibronectin deposition; Focal adhesion kinase; P190RhoGAP; RhoA Contractility.
Copyright © 2021 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Figures
Similar articles
-
Fibronectin-dependent integrin signaling drives EphA2 expression in vascular smooth muscle cells.Am J Physiol Cell Physiol. 2025 May 1;328(5):C1623-C1636. doi: 10.1152/ajpcell.01021.2024. Epub 2025 Apr 16. Am J Physiol Cell Physiol. 2025. PMID: 40241381
-
The fibronectin-binding integrins alpha5beta1 and alphavbeta3 differentially modulate RhoA-GTP loading, organization of cell matrix adhesions, and fibronectin fibrillogenesis.J Cell Biol. 2002 Dec 23;159(6):1071-86. doi: 10.1083/jcb.200205014. Epub 2002 Dec 16. J Cell Biol. 2002. PMID: 12486108 Free PMC article.
-
Characterization of stitch adhesions: Fibronectin-containing cell-cell contacts formed by fibroblasts.Exp Cell Res. 2019 Nov 1;384(1):111616. doi: 10.1016/j.yexcr.2019.111616. Epub 2019 Sep 6. Exp Cell Res. 2019. PMID: 31499058 Free PMC article.
-
Integrins in cell adhesion and signaling.Hum Cell. 1996 Sep;9(3):181-6. Hum Cell. 1996. PMID: 9183647 Review.
-
Tensins: Bridging AMP-Activated Protein Kinase with Integrin Activation.Trends Cell Biol. 2017 Oct;27(10):703-711. doi: 10.1016/j.tcb.2017.06.004. Epub 2017 Jul 8. Trends Cell Biol. 2017. PMID: 28698049 Review.
Cited by
-
Eph receptor signaling complexes in the plasma membrane.Trends Biochem Sci. 2024 Dec;49(12):1079-1096. doi: 10.1016/j.tibs.2024.10.002. Epub 2024 Nov 12. Trends Biochem Sci. 2024. PMID: 39537538 Review.
-
Inhibition of EphA2 by syndecan-4 in wounded skin regulates clustering of fibroblasts.J Mol Cell Biol. 2025 Jun 28;17(1):mjae054. doi: 10.1093/jmcb/mjae054. J Mol Cell Biol. 2025. PMID: 39716457 Free PMC article.
-
CK2β Is a Gatekeeper of Focal Adhesions Regulating Cell Spreading.Front Mol Biosci. 2022 Jun 29;9:900947. doi: 10.3389/fmolb.2022.900947. eCollection 2022. Front Mol Biosci. 2022. PMID: 35847979 Free PMC article.
-
EphrinA5 regulates cell motility by modulating Snhg15/DNA triplex-dependent targeting of DNMT1 to the Ncam1 promoter.Epigenetics Chromatin. 2023 Oct 26;16(1):42. doi: 10.1186/s13072-023-00516-4. Epigenetics Chromatin. 2023. PMID: 37880732 Free PMC article.
-
Eph receptors and ephrins in cancer progression.Nat Rev Cancer. 2024 Jan;24(1):5-27. doi: 10.1038/s41568-023-00634-x. Epub 2023 Nov 23. Nat Rev Cancer. 2024. PMID: 37996538 Free PMC article. Review.
References
-
- Lock JG, Wehrle-Haller B, Stromblad S, Cell-matrix adhesion complexes: master control machinery of cell migration, Seminars in cancer biology 18(1) (2008) 65–76. - PubMed
-
- Valiente-Alandi I, Potter SJ, Salvador AM, Schafer AE, Schips T, Carrillo-Salinas F, Gibson AM, Nieman ML, Perkins C, Sargent MA, Huo J, Lorenz JN, DeFalco T, Molkentin JD, Alcaide P, Blaxall BC, Inhibiting Fibronectin Attenuates Fibrosis and Improves Cardiac Function in a Model of Heart Failure, Circulation 138(12) (2018) 1236–1252. - PMC - PubMed
-
- Wang WY, Twu CW, Liu YC, Lin HH, Chen CJ, Lin JC, Fibronectin promotes nasopharyngeal cancer cell motility and proliferation, Biomed Pharmacother 109 (2019) 1772–1784. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
