

Colicin-Mediated Transport of DNA through the Iron Transporter FepA
- PMID: 34544275
- PMCID: PMC8546555
- DOI: 10.1128/mBio.01787-21
Colicin-Mediated Transport of DNA through the Iron Transporter FepA
Abstract
Colicins are protein antibiotics deployed by Escherichia coli to eliminate competing strains. Colicins frequently exploit outer membrane (OM) nutrient transporters to penetrate the selectively permeable bacterial cell envelope. Here, by applying live-cell fluorescence imaging, we were able to monitor the entry of the pore-forming toxin colicin B (ColB) into E. coli and localize it within the periplasm. We further demonstrate that single-stranded DNA coupled to ColB can also be transported to the periplasm, emphasizing that the import routes of colicins can be exploited to carry large cargo molecules into bacteria. Moreover, we characterize the molecular mechanism of ColB association with its OM receptor FepA by applying a combination of photoactivated cross-linking, mass spectrometry, and structural modeling. We demonstrate that complex formation is coincident with large-scale conformational changes in the colicin. Thereafter, active transport of ColB through FepA involves the colicin taking the place of the N-terminal half of the plug domain that normally occludes this iron transporter. IMPORTANCE Decades of excessive use of readily available antibiotics has generated a global problem of antibiotic resistance and, hence, an urgent need for novel antibiotic solutions. Bacteriocins are protein-based antibiotics produced by bacteria to eliminate closely related competing bacterial strains. Bacteriocin toxins have evolved to bypass the complex cell envelope in order to kill bacterial cells. Here, we uncover the cellular penetration mechanism of a well-known but poorly understood bacteriocin called colicin B that is active against Escherichia coli. Moreover, we demonstrate that the colicin B-import pathway can be exploited to deliver conjugated DNA cargo into bacterial cells. Our work leads to a better understanding of the way bacteriocins, as potential alternative antibiotics, execute their mode of action as well as highlighting how they might even be exploited in the genomic manipulation of Gram-negative bacteria.
Keywords: DNA delivery; Rosetta flexible backbone; antibiotic resistance; bacteriocins; conformational changes; membrane transport.
Figures
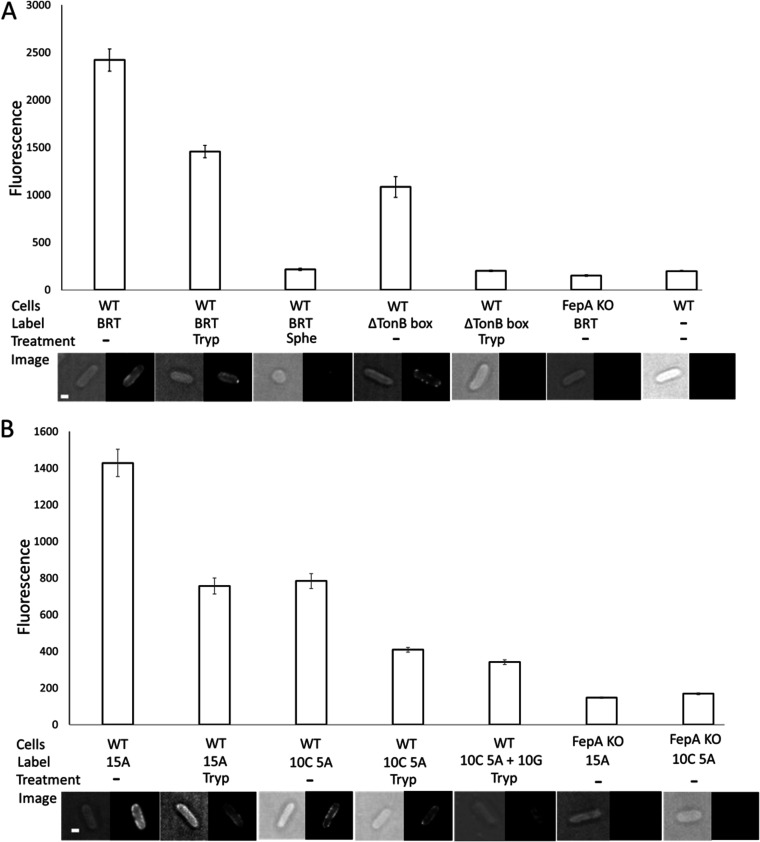
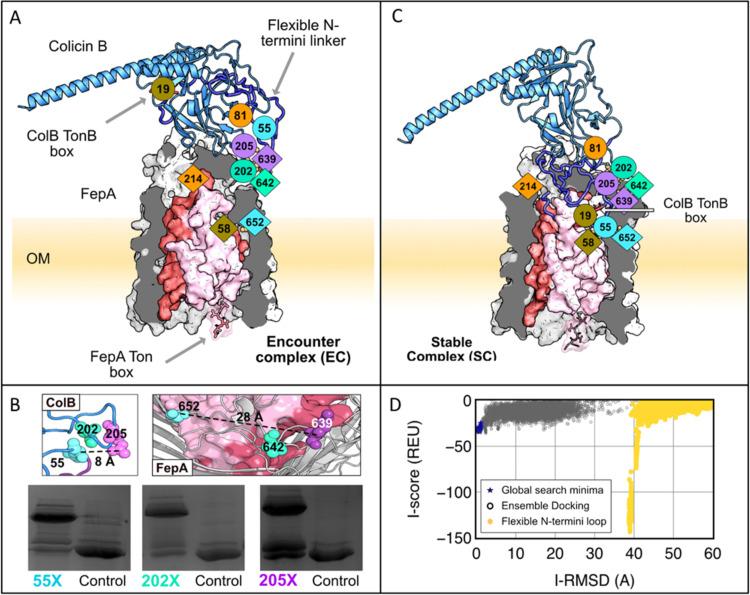
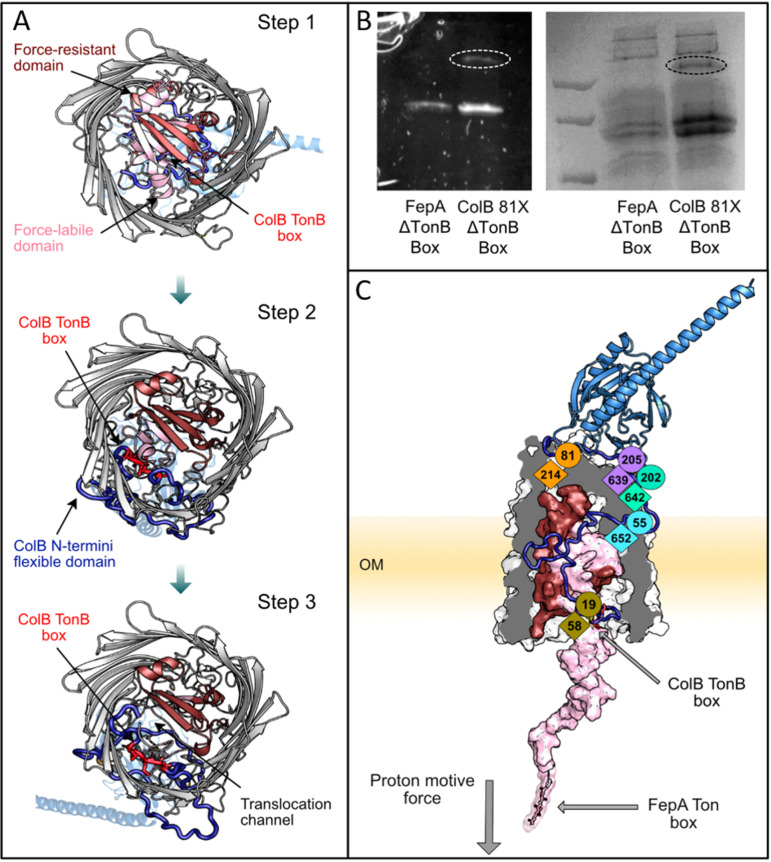
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
