

RNA m6A demethylase FTO-mediated epigenetic up-regulation of LINC00022 promotes tumorigenesis in esophageal squamous cell carcinoma
- PMID: 34544449
- PMCID: PMC8451109
- DOI: 10.1186/s13046-021-02096-1
RNA m6A demethylase FTO-mediated epigenetic up-regulation of LINC00022 promotes tumorigenesis in esophageal squamous cell carcinoma
Abstract
Background: Long non-coding RNA (LncRNA) controls cell proliferation and plays a significant role in the initiation and progression of esophageal squamous cell carcinoma (ESCC). N6-methyladenosine (m6A) modification now is recognized as a master driver of RNA function to maintain homeostasis in cancer cells. However, how m6A regulates LncRNA function and its role in tumorigenesis of ESCC remain unclear.
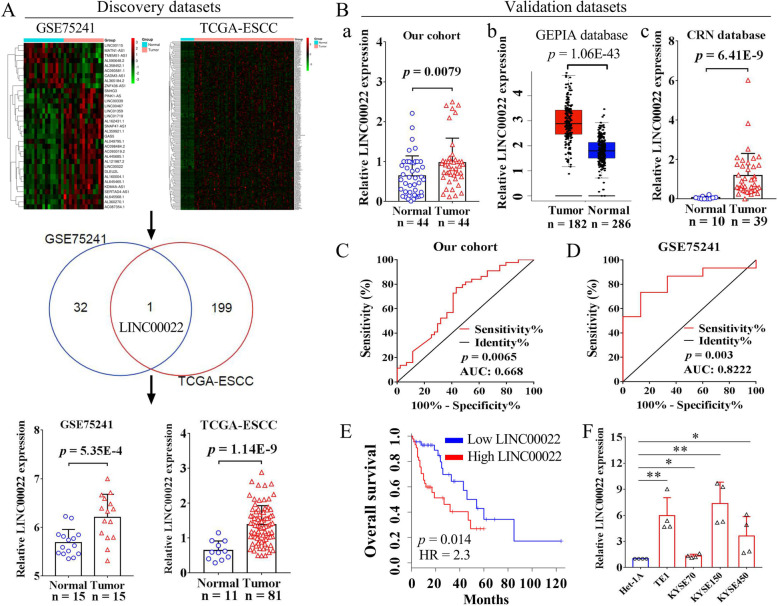
Methods: Multiple ESCC datasets were used to analyze gene expression in tumor tissues and normal tissues. Kaplan-Meier method and the ROC curve were conducted to evaluate the prognostic value and diagnostic value of LINC00022 in ESCC, respectively. Both gain-of-function and loss-of-function experiments were employed to investigate the effects of LINC00022 on ESCC growth in vitro and in vivo. Bioinformatics analysis, colorimetric m6A assay, RIP, MeRIP and co-IP was performed to explore the epigenetic mechanism of LINC00022 up-regulation in ESCC.
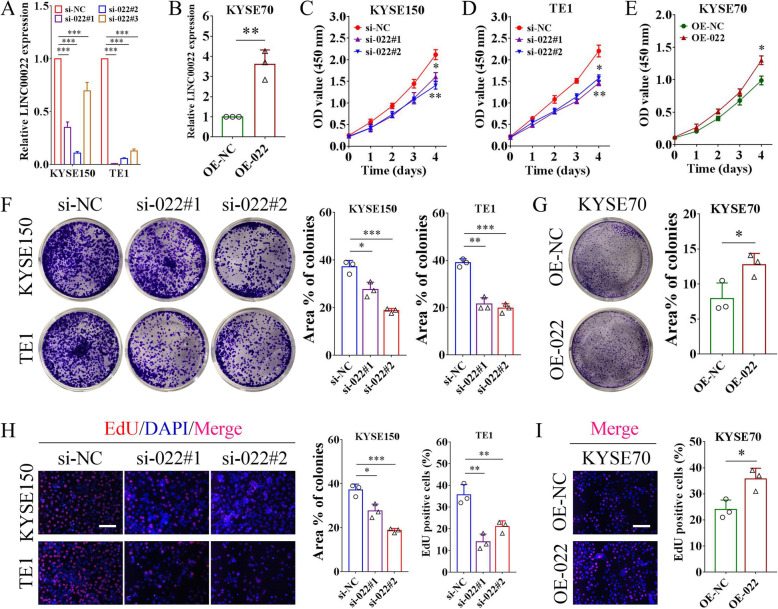
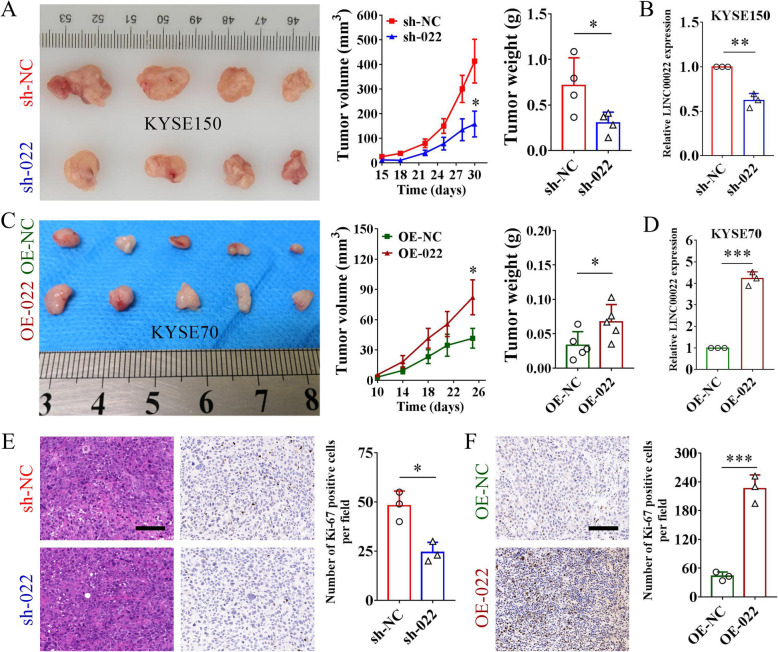
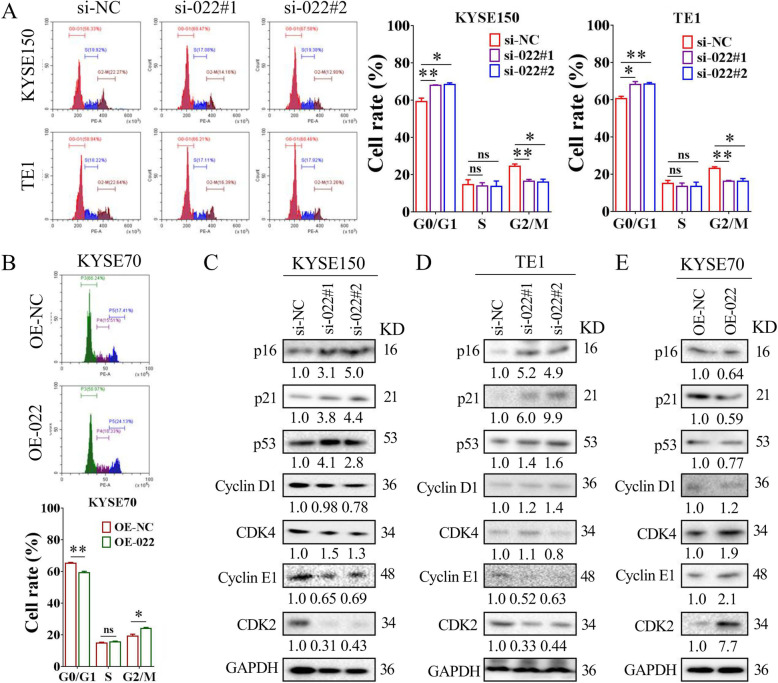
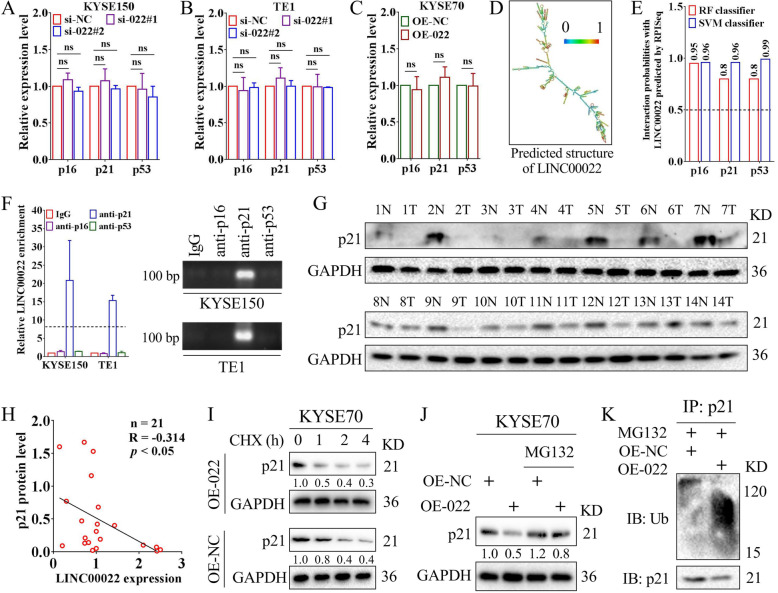
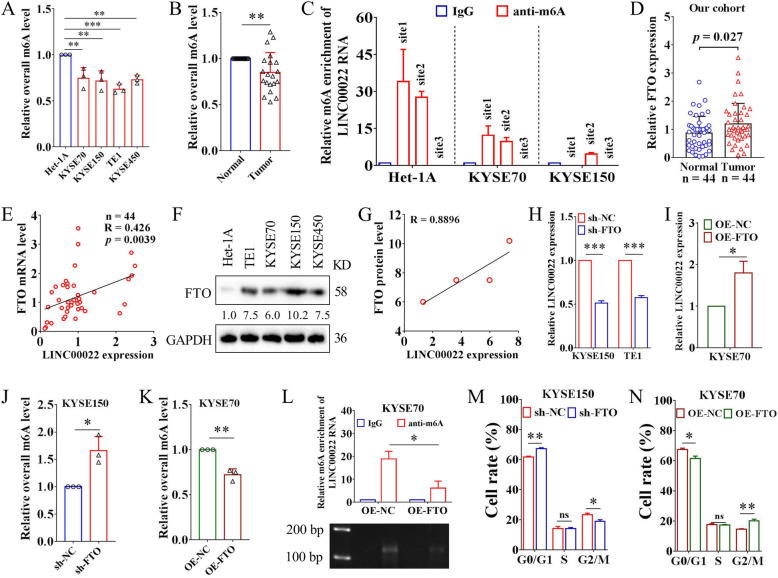
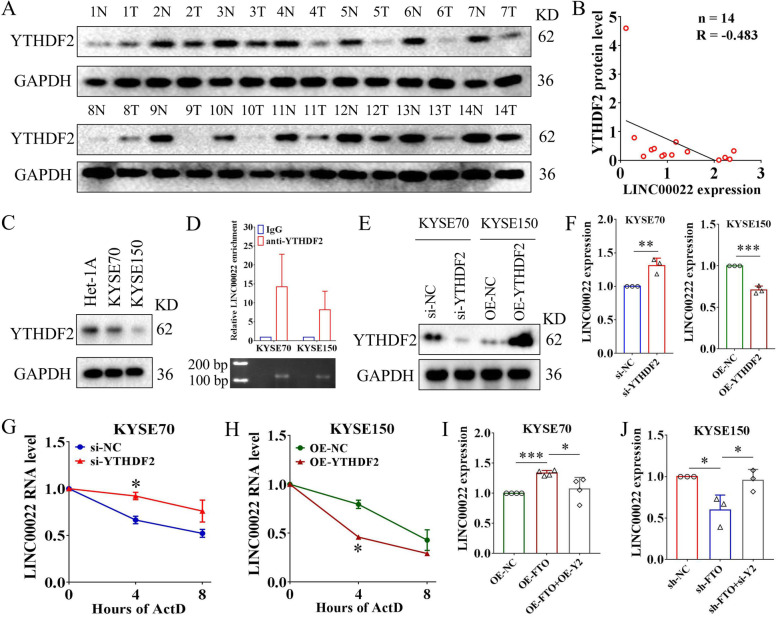
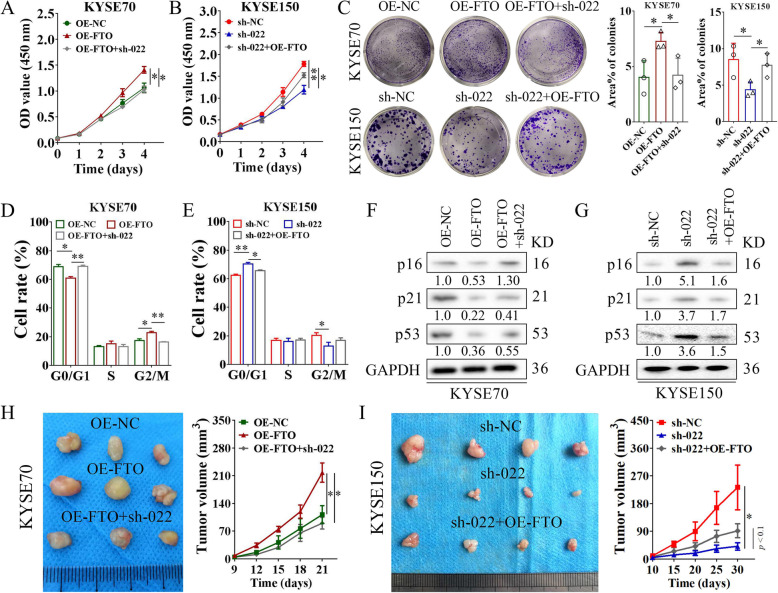
Results: Here we report that m6A demethylation of LncRNA LINC00022 by fat mass and obesity-associated protein (FTO) promotes tumor growth of ESCC in vivo. Clinically, we revealed that LINC00022 was up-regulated in primary ESCC samples and was predictive of poor clinical outcome for ESCC patients. Mechanistically, LINC00022 directly binds to p21 protein and promotes its ubiquitination-mediated degradation, thereby facilitating cell-cycle progression and proliferation. Further, the elevated FTO in ESCC decreased m6A methylation of LINC00022 transcript, leading to the inhibition of LINC00022 decay via the m6A reader YTHDF2. Over-expression of FTO was shown to drive LINC00022-dependent cell proliferation and tumor growth of ESCC.
Conclusions: Thus, this study demonstrated m6A-mediated epigenetic modification of LncRNA contributes to the tumorigenesis in ESCC and LINC00022, specific target of m6A, serves as a potential biomarker for this malignancy.
Keywords: Cell cycle; Esophageal squamous cell carcinoma; FTO; LINC00022; N6-methyladenosine; Tumorigenesis.
© 2021. The Author(s).
Conflict of interest statement
No potential conflicts of interest.
Figures
References
MeSH terms
Substances
Grants and funding
- U2004201/key programme
- SBGJ202003053/science and technology department, henan province
- LHGJ20191054/science and technology department of henan province
- LHGJ20200765, SB201903032/science and technology department of henan province
- XKZDJC202001/Key Discipline Construction Project for Prevention and Treatment of Esophageal Cancer in Zhengzhou University
LinkOut - more resources
Full Text Sources
Miscellaneous
