

Identification of limb-specific Lmx1b auto-regulatory modules with Nail-patella syndrome pathogenicity
- PMID: 34545091
- PMCID: PMC8452625
- DOI: 10.1038/s41467-021-25844-5
Identification of limb-specific Lmx1b auto-regulatory modules with Nail-patella syndrome pathogenicity
Abstract
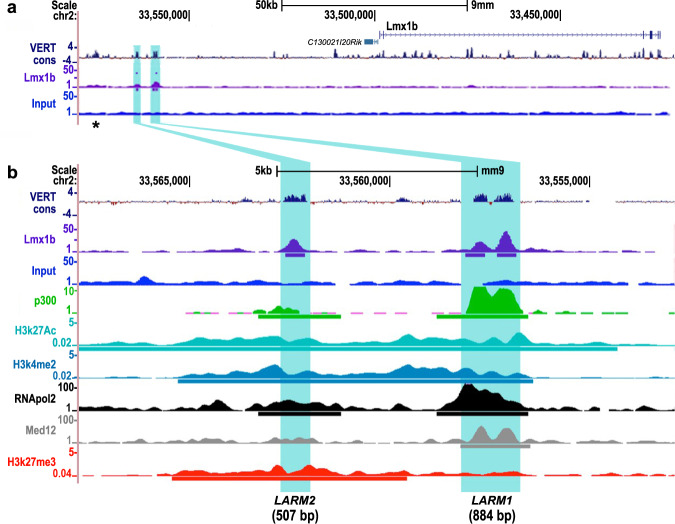
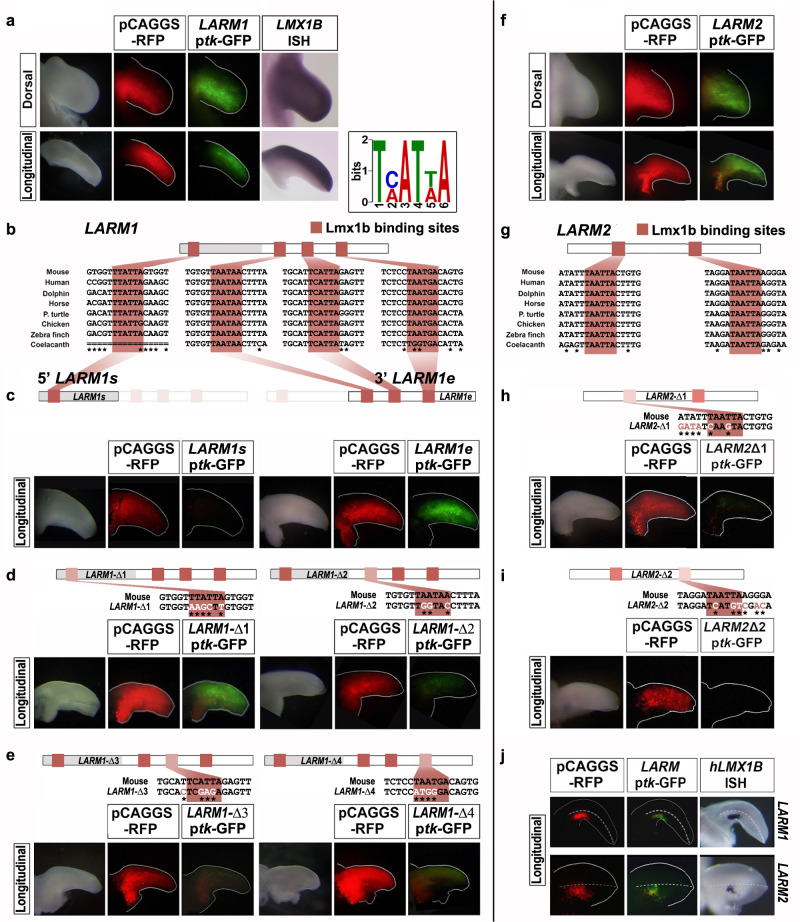
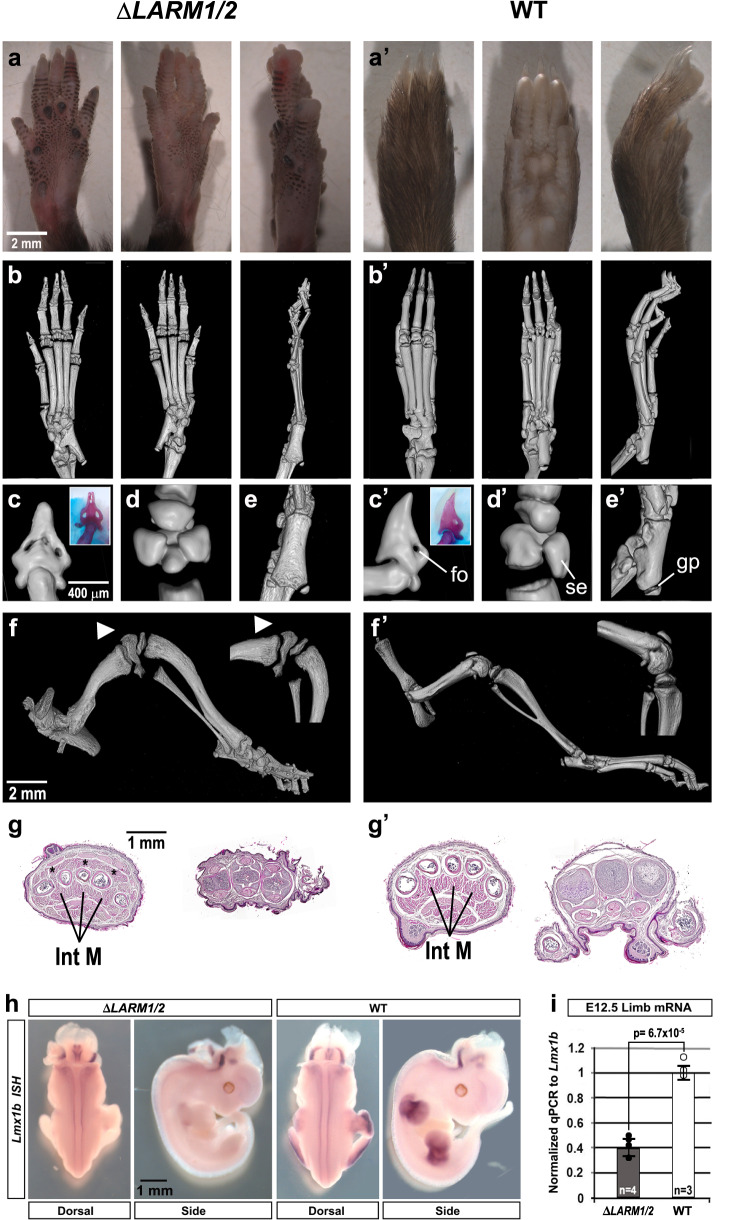
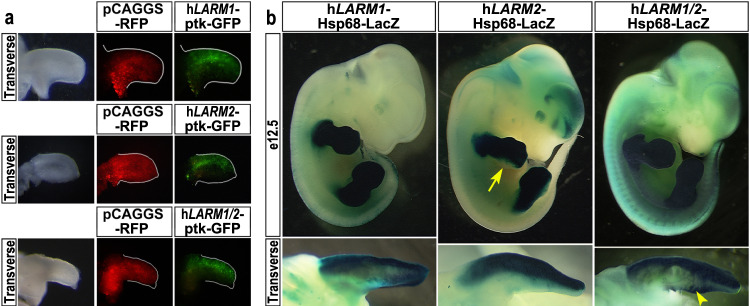
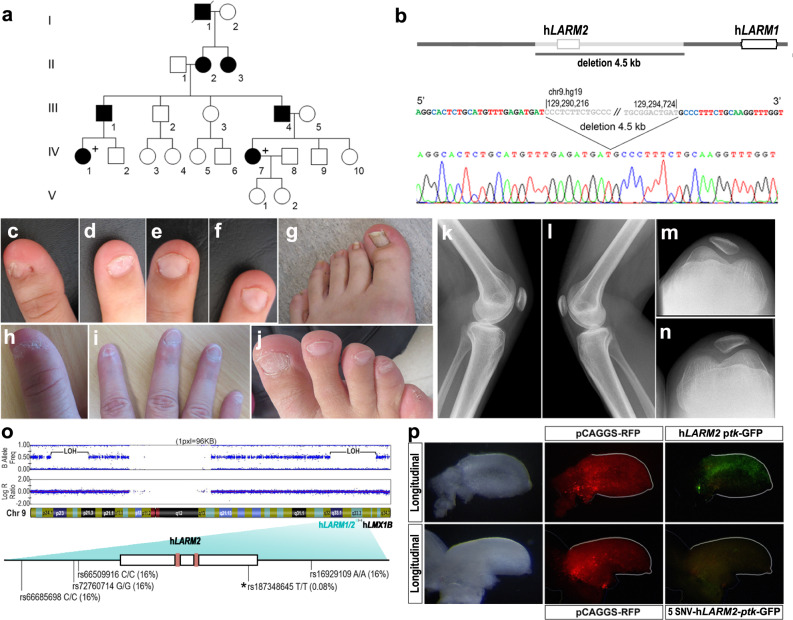
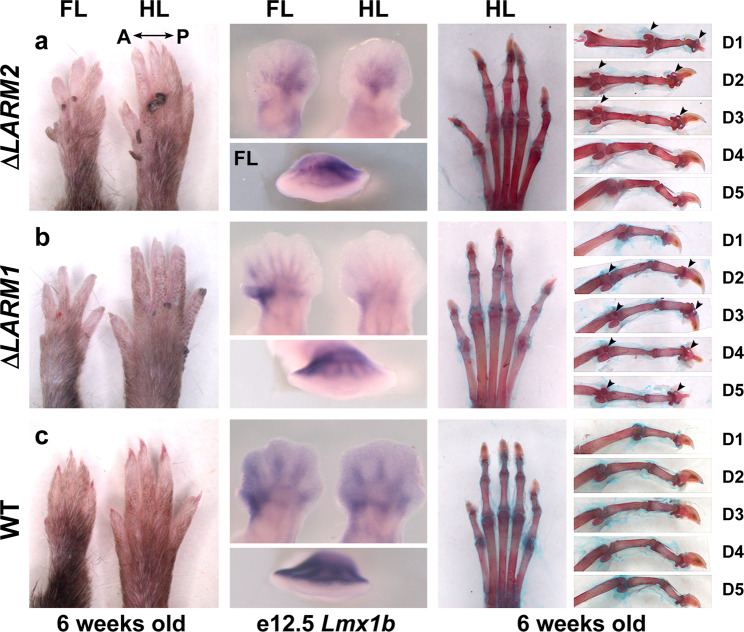
LMX1B haploinsufficiency causes Nail-patella syndrome (NPS; MIM 161200), characterized by nail dysplasia, absent/hypoplastic patellae, chronic kidney disease, and glaucoma. Accordingly in mice, Lmx1b has been shown to play crucial roles in the development of the limb, kidney and eye. Although one functional allele of Lmx1b appears adequate for development, Lmx1b null mice display ventral-ventral distal limbs with abnormal kidney, eye and cerebellar development, more disruptive, but fully concordant with NPS. In Lmx1b functional knockouts (KOs), Lmx1b transcription in the limb is decreased nearly 6-fold, indicating autoregulation. Herein, we report on two conserved Lmx1b-associated cis-regulatory modules (LARM1 and LARM2) that are bound by Lmx1b, amplify Lmx1b expression with unique spatial modularity in the limb, and are necessary for Lmx1b-mediated limb dorsalization. These enhancers, being conserved across vertebrates (including coelacanth, but not other fish species), and required for normal locomotion, provide a unique opportunity to study the role of dorsalization in the fin to limb transition. We also report on two NPS patient families with normal LMX1B coding sequence, but with loss-of-function variations in the LARM1/2 region, stressing the role of regulatory modules in disease pathogenesis.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
