

Mechanisms governing activity-dependent synaptic pruning in the developing mammalian CNS
- PMID: 34545240
- PMCID: PMC8541743
- DOI: 10.1038/s41583-021-00507-y
Mechanisms governing activity-dependent synaptic pruning in the developing mammalian CNS
Abstract
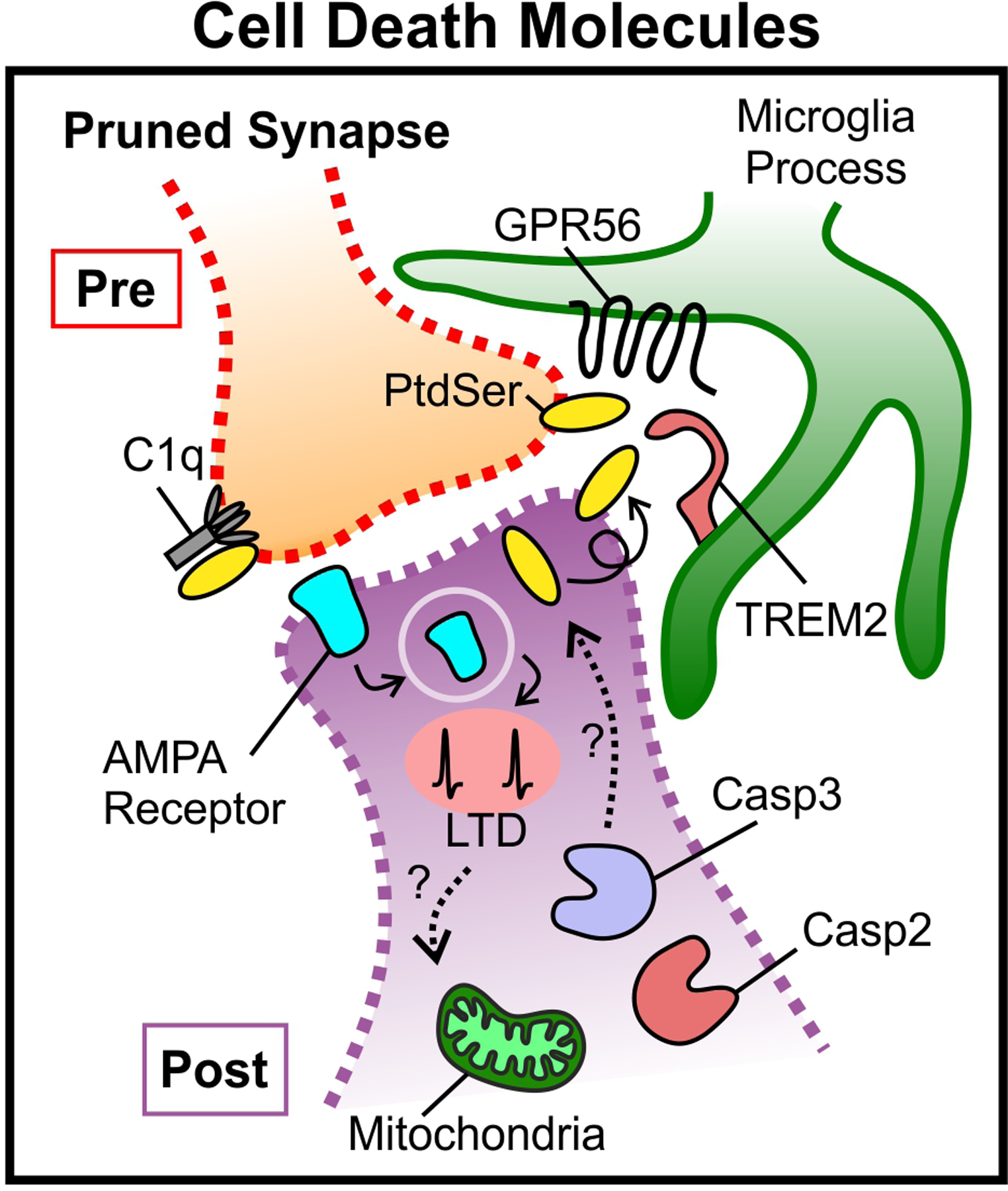
Almost 60 years have passed since the initial discovery by Hubel and Wiesel that changes in neuronal activity can elicit developmental rewiring of the central nervous system (CNS). Over this period, we have gained a more comprehensive picture of how both spontaneous neural activity and sensory experience-induced changes in neuronal activity guide CNS circuit development. Here we review activity-dependent synaptic pruning in the mammalian CNS, which we define as the removal of a subset of synapses, while others are maintained, in response to changes in neural activity in the developing nervous system. We discuss the mounting evidence that immune and cell-death molecules are important mechanistic links by which changes in neural activity guide the pruning of specific synapses, emphasizing the role of glial cells in this process. Finally, we discuss how these developmental pruning programmes may go awry in neurodevelopmental disorders of the human CNS, focusing on autism spectrum disorder and schizophrenia. Together, our aim is to give an overview of how the field of activity-dependent pruning research has evolved, led to exciting new questions and guided the identification of new, therapeutically relevant mechanisms that result in aberrant circuit development in neurodevelopmental disorders.
© 2021. Springer Nature Limited.
Conflict of interest statement
Figures



References
-
- Sanes JR & Lichtman JW Development of the vertebrate neuromuscular junction. Annual review of neuroscience 22, 389–442 (1999). - PubMed
