

Metabolic channeling: predictions, deductions, and evidence
- PMID: 34547238
- PMCID: PMC8485759
- DOI: 10.1016/j.molcel.2021.08.030
Metabolic channeling: predictions, deductions, and evidence
Abstract
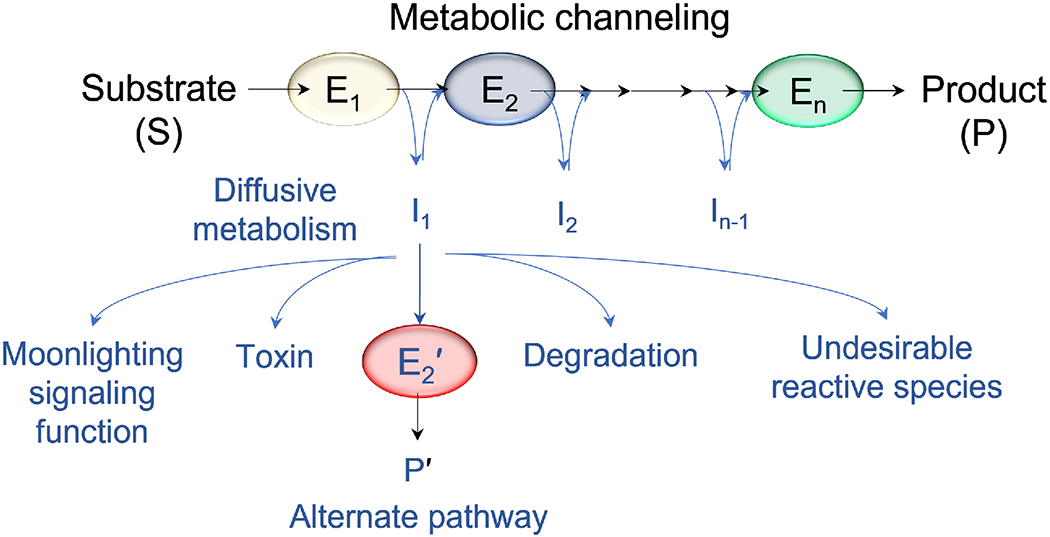
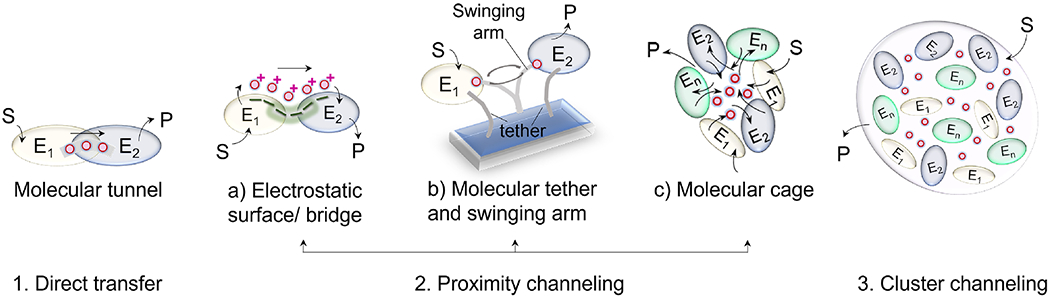
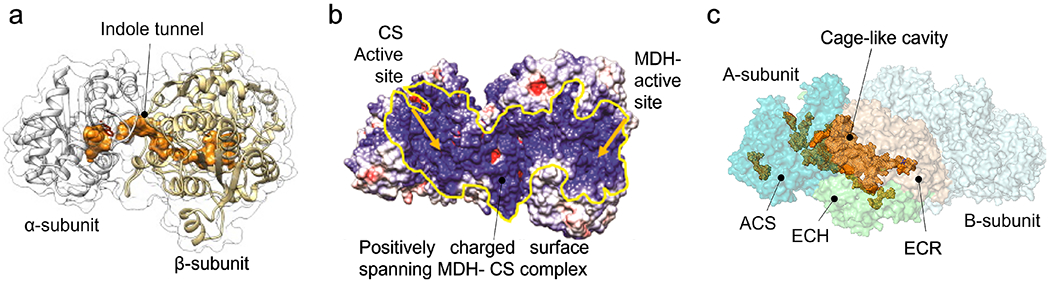
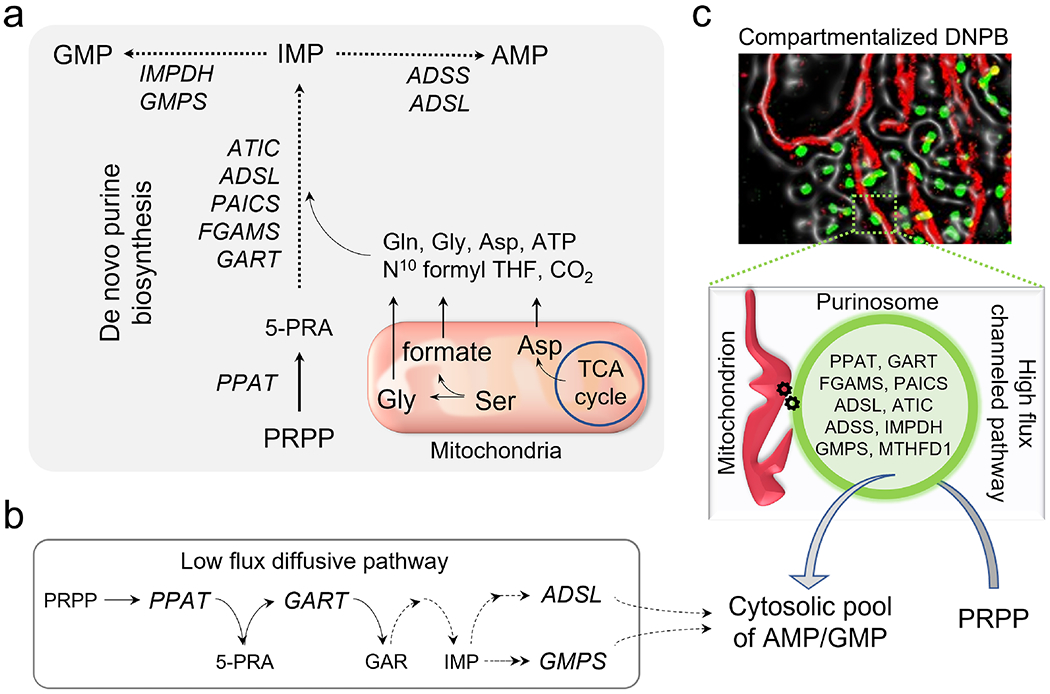
With the elucidation of myriad anabolic and catabolic enzyme-catalyzed cellular pathways crisscrossing each other, an obvious question arose: how could these networks operate with maximal catalytic efficiency and minimal interference? A logical answer was the postulate of metabolic channeling, which in its simplest embodiment assumes that the product generated by one enzyme passes directly to a second without diffusion into the surrounding medium. This tight coupling of activities might increase a pathway's metabolic flux and/or serve to sequester unstable/toxic/reactive intermediates as well as prevent their access to other networks. Here, we present evidence for this concept, commencing with enzymes that feature a physical molecular tunnel, to multi-enzyme complexes that retain pathway substrates through electrostatics or enclosures, and finally to metabolons that feature collections of enzymes assembled into clusters with variable stoichiometric composition. Lastly, we discuss the advantages of reversibly assembled metabolons in the context of the purinosome, the purine biosynthesis metabolon.
Keywords: membrane-less compartmentalization; metabolic channeling; metabolon; molecular tunnel; purinosome.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Ahmed SA, Ruvinov SB, Kayastha AM, and Miles EW (1991). Mechanism of mutual activation of the tryptophan synthase alpha and beta subunits. Analysis of the reaction specificity and substrate-induced inactivation of active site and tunnel mutants of the beta subunit. J Biol Chem 266, 21548–21557. - PubMed
-
- Alber BE, and Fuchs G (2002). Propionyl-coenzyme A synthase from Chloroflexus aurantiacus, a key enzyme of the 3-hydroxypropionate cycle for autotrophic CO2 fixation. J Biol Chem 277, 12137–12143. - PubMed
-
- Amaro RE, Sethi A, Myers RS, Davisson VJ, and Luthey-Schulten ZA (2007). A Network of Conserved Interactions Regulates the Allosteric Signal in a Glutamine Amidotransferase. Biochemistry 46, 2156–2173. - PubMed
-
- Anderson KS, Kim AY, Quillen JM, Sayers E, Yang X-J, and Miles EW (1995). Kinetic Characterization of Channel Impaired Mutants of Tryptophan Synthase (*). Journal of Biological Chemistry 270, 29936–29944. - PubMed
-
- Anderson KS, Miles EW, and Johnson KA (1991). Serine modulates substrate channeling in tryptophan synthase. A novel intersubunit triggering mechanism. J Biol Chem 266, 8020–8033. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
