

Assembly and organization of the N-terminal region of mucin MUC5AC: Indications for structural and functional distinction from MUC5B
- PMID: 34548396
- PMCID: PMC8488587
- DOI: 10.1073/pnas.2104490118
Assembly and organization of the N-terminal region of mucin MUC5AC: Indications for structural and functional distinction from MUC5B
Abstract
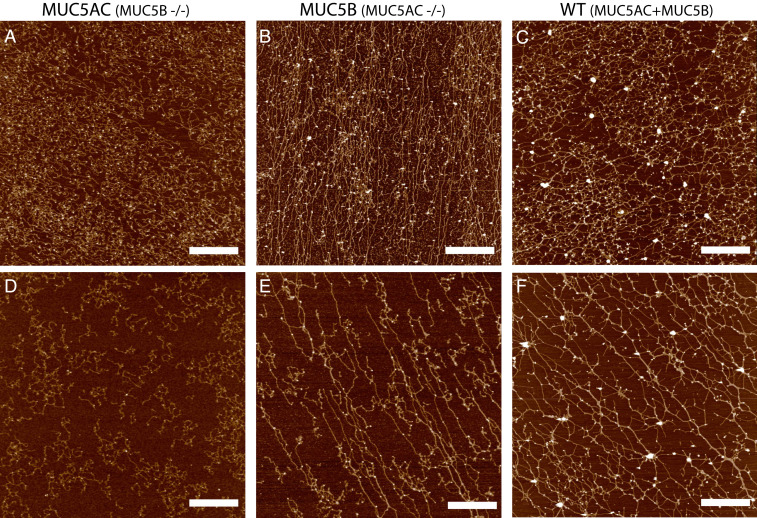
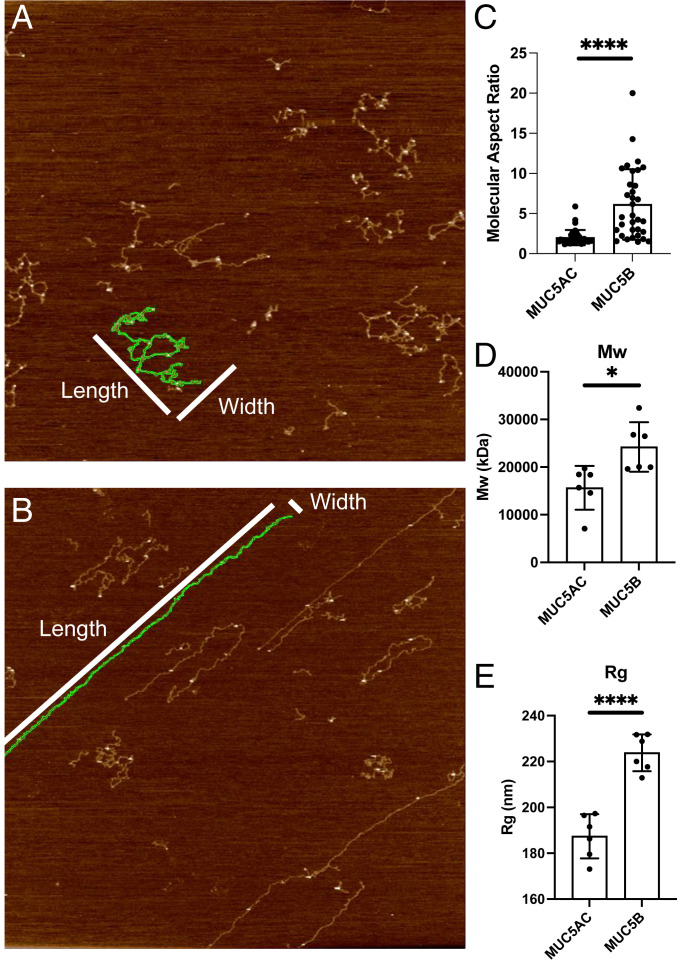
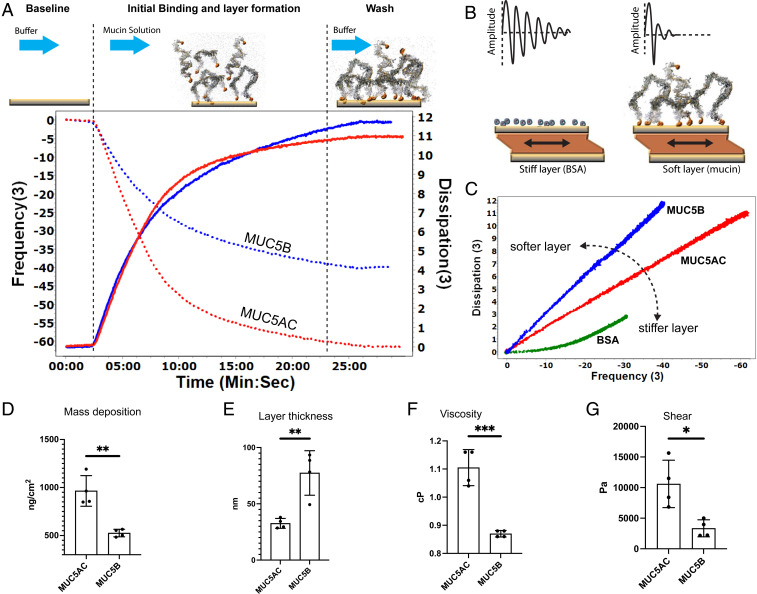
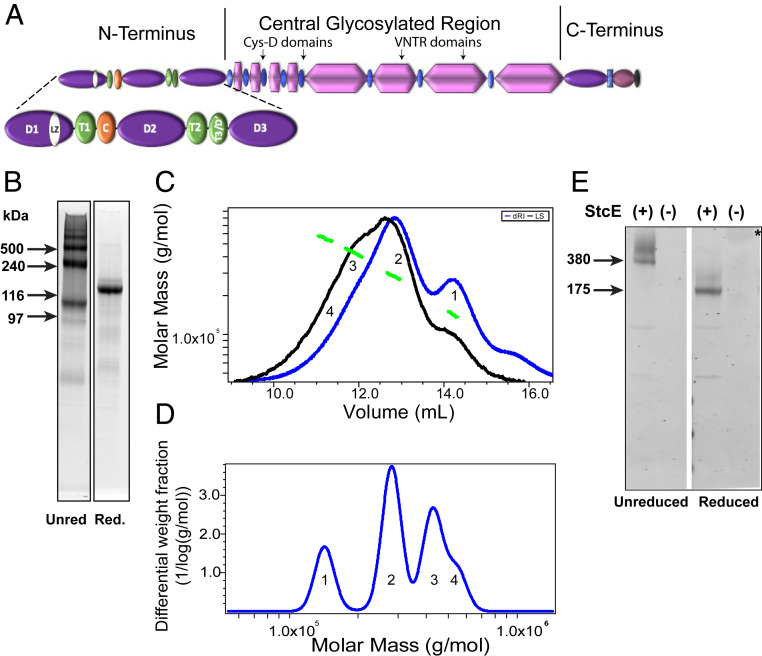
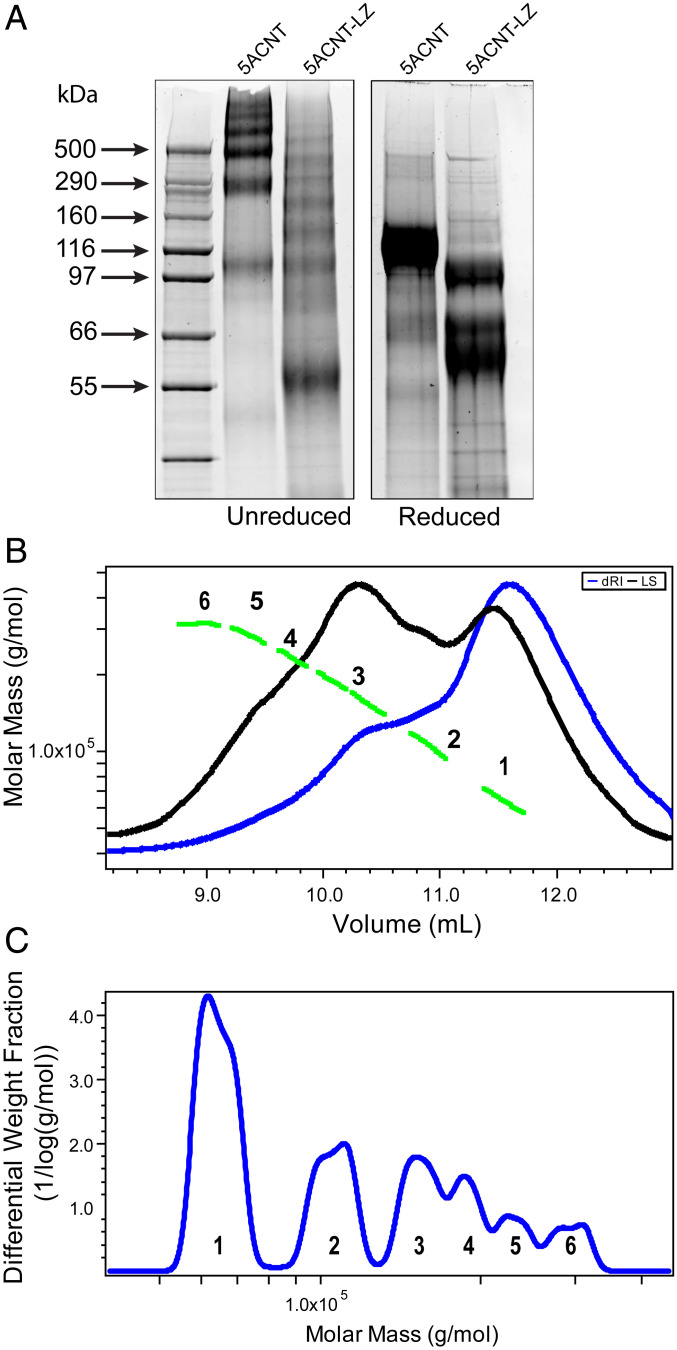
Elevated levels of MUC5AC, one of the major gel-forming mucins in the lungs, are closely associated with chronic obstructive lung diseases such as chronic bronchitis and asthma. It is not known, however, how the structure and/or gel-making properties of MUC5AC contribute to innate lung defense in health and drive the formation of stagnant mucus in disease. To understand this, here we studied the biophysical properties and macromolecular assembly of MUC5AC compared to MUC5B. To study each native mucin, we used Calu3 monomucin cultures that produced MUC5AC or MUC5B. To understand the macromolecular assembly of MUC5AC through N-terminal oligomerization, we expressed a recombinant whole N-terminal domain (5ACNT). Scanning electron microscopy and atomic force microscopy imaging indicated that the two mucins formed distinct networks on epithelial and experimental surfaces; MUC5B formed linear, infrequently branched multimers, whereas MUC5AC formed tightly organized networks with a high degree of branching. Quartz crystal microbalance-dissipation monitoring experiments indicated that MUC5AC bound significantly more to hydrophobic surfaces and was stiffer and more viscoelastic as compared to MUC5B. Light scattering analysis determined that 5ACNT primarily forms disulfide-linked covalent dimers and higher-order oligomers (i.e., trimers and tetramers). Selective proteolytic digestion of the central glycosylated region of the full-length molecule confirmed that MUC5AC forms dimers and higher-order oligomers through its N terminus. Collectively, the distinct N-terminal organization of MUC5AC may explain the more adhesive and unique viscoelastic properties of branched, highly networked MUC5AC gels. These properties may generate insight into why/how MUC5AC forms a static, "tethered" mucus layer in chronic muco-obstructive lung diseases.
Keywords: MUC5AC; airways; lung; mucin; mucus gel.
Conflict of interest statement
The authors declare no competing interest.
Figures

References
-
- Veerman E. C., et al., Distinct localization of MUC5B glycoforms in the human salivary glands. Glycobiology 13, 363–366 (2003). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
