Spatial regulation by multiple Gremlin1 enhancers provides digit development with cis-regulatory robustness and evolutionary plasticity
- PMID: 34548488
- PMCID: PMC8455560
- DOI: 10.1038/s41467-021-25810-1
Spatial regulation by multiple Gremlin1 enhancers provides digit development with cis-regulatory robustness and evolutionary plasticity
Abstract
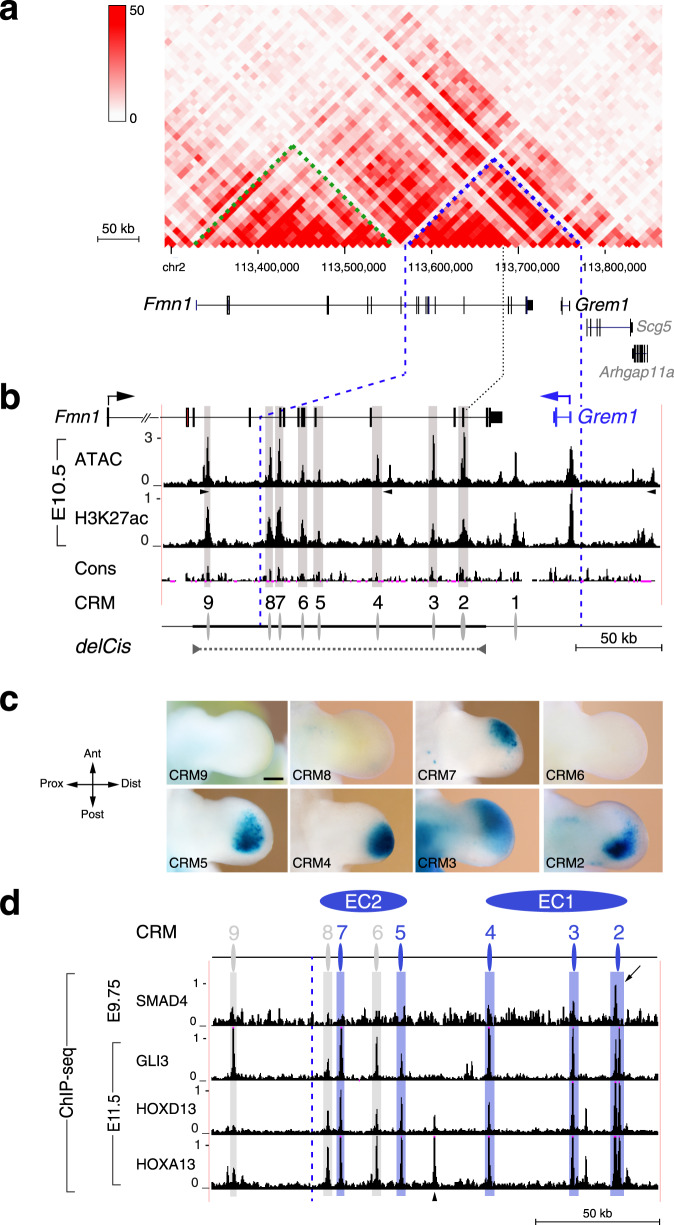
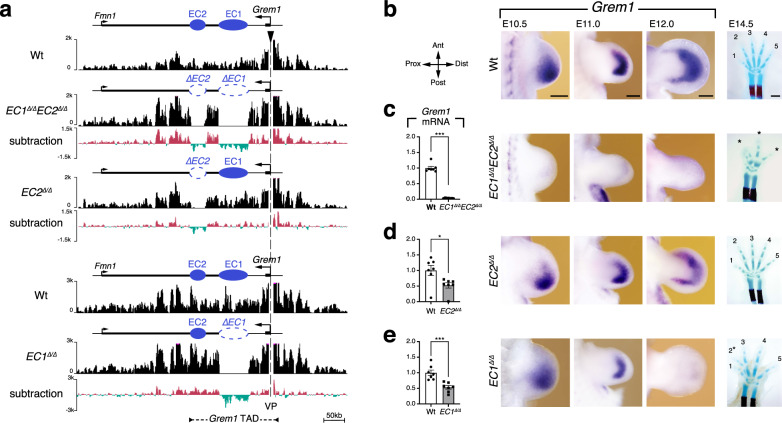
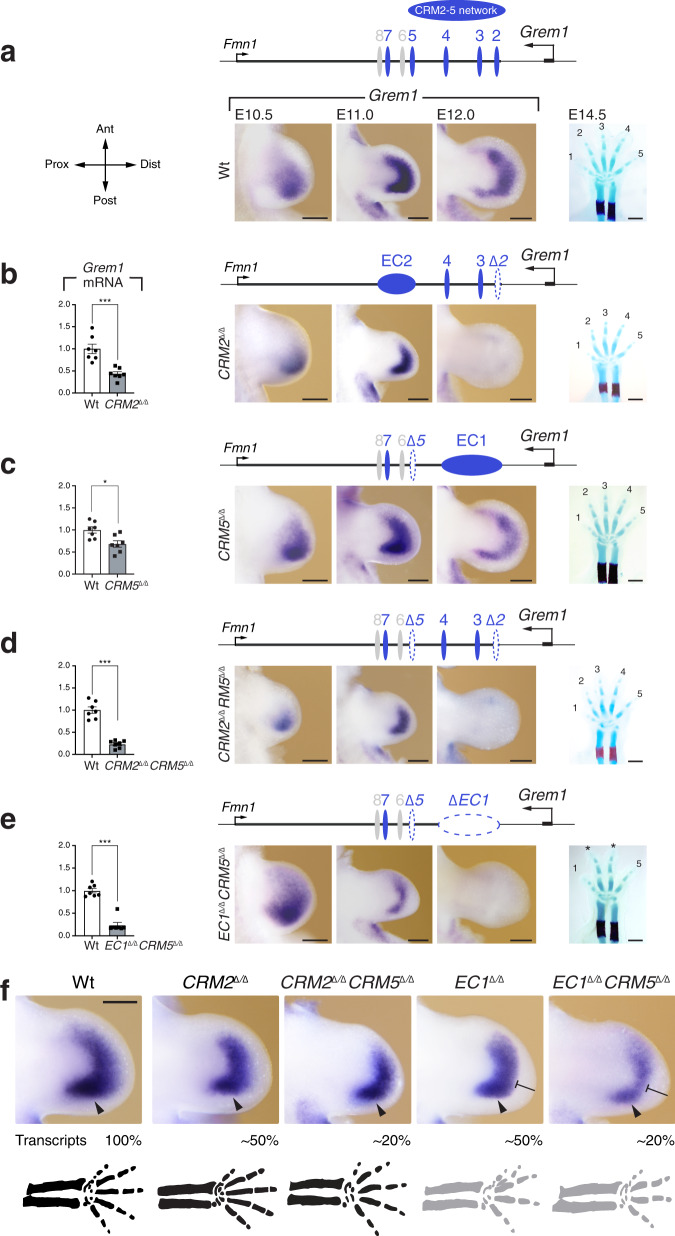
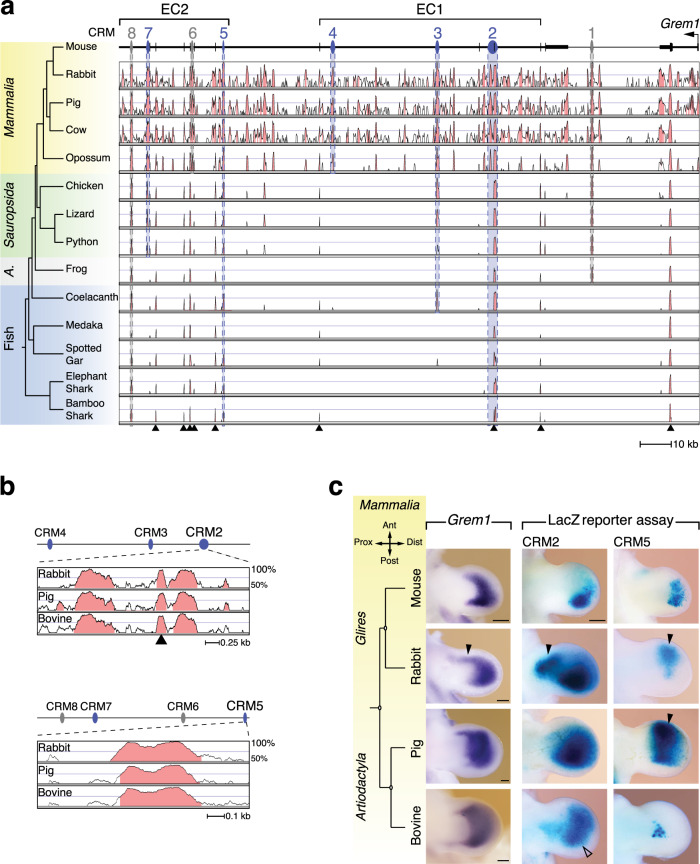
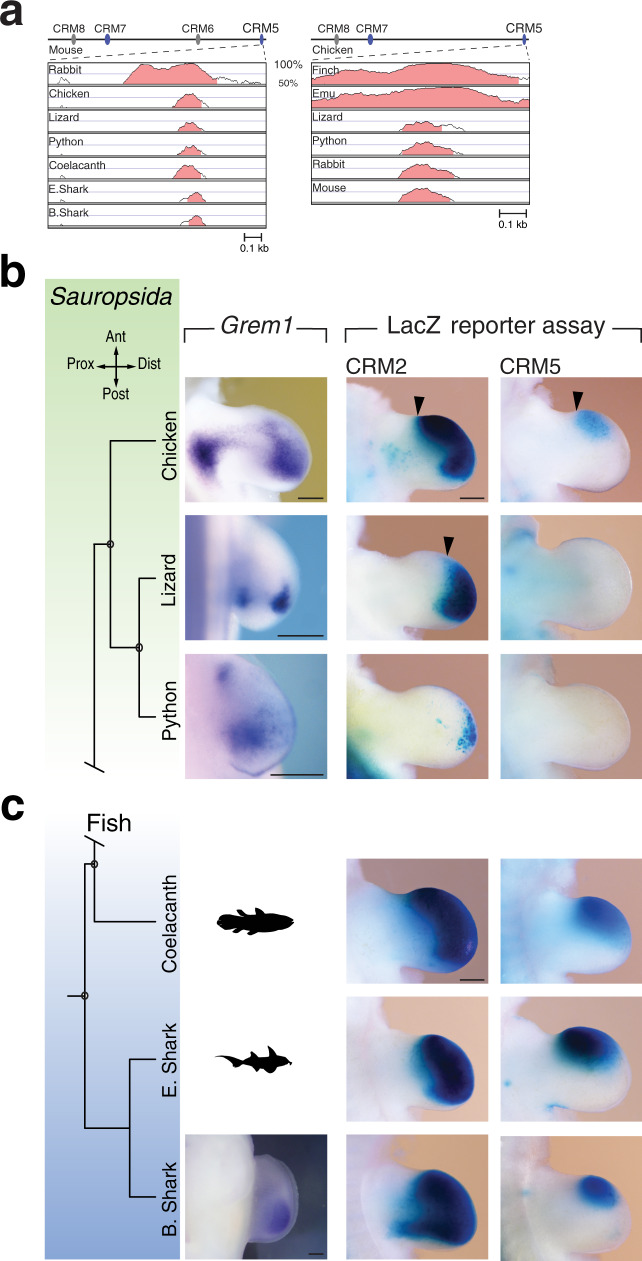
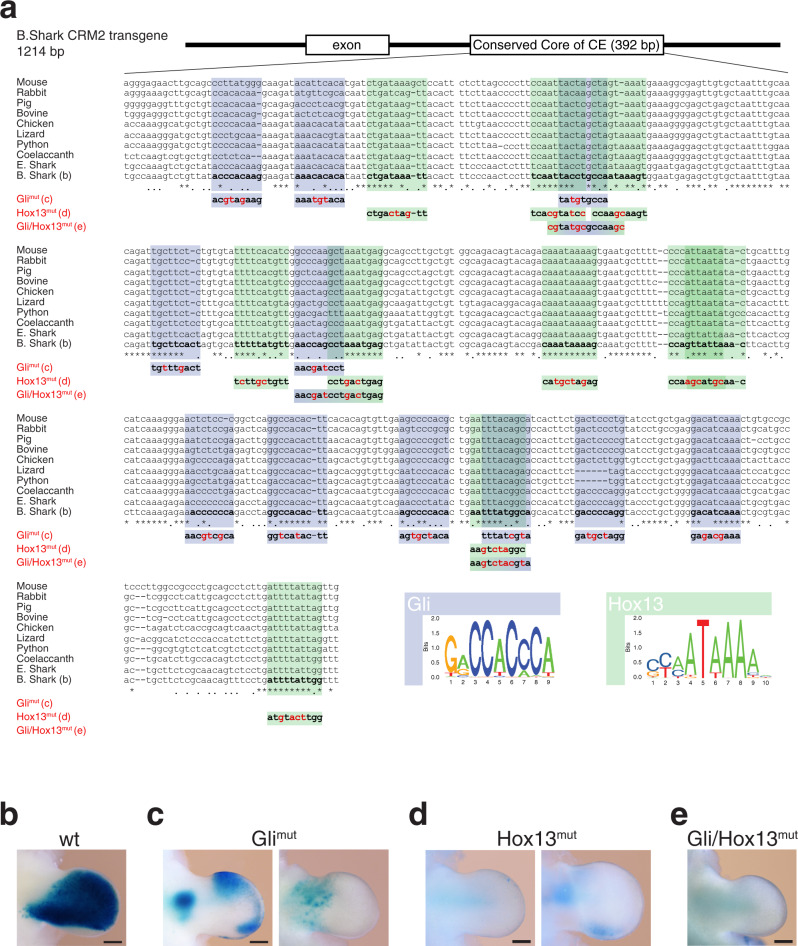
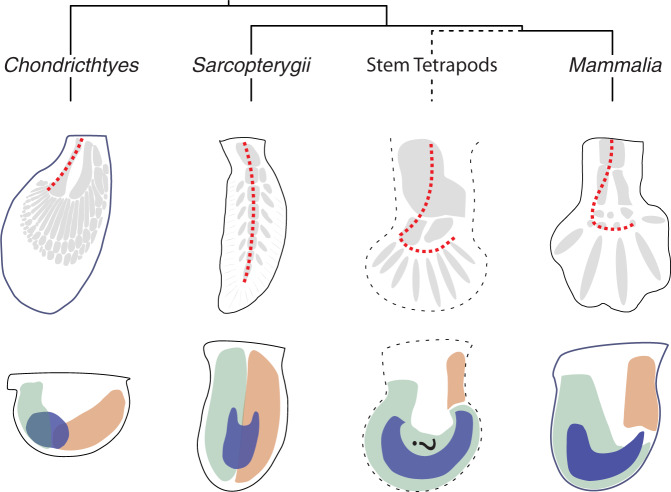
Precise cis-regulatory control of gene expression is essential for normal embryogenesis and tissue development. The BMP antagonist Gremlin1 (Grem1) is a key node in the signalling system that coordinately controls limb bud development. Here, we use mouse reverse genetics to identify the enhancers in the Grem1 genomic landscape and the underlying cis-regulatory logics that orchestrate the spatio-temporal Grem1 expression dynamics during limb bud development. We establish that transcript levels are controlled in an additive manner while spatial regulation requires synergistic interactions among multiple enhancers. Disrupting these interactions shows that altered spatial regulation rather than reduced Grem1 transcript levels prefigures digit fusions and loss. Two of the enhancers are evolutionary ancient and highly conserved from basal fishes to mammals. Analysing these enhancers from different species reveal the substantial spatial plasticity in Grem1 regulation in tetrapods and basal fishes, which provides insights into the fin-to-limb transition and evolutionary diversification of pentadactyl limbs.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures