

Integration of feeding behavior by the liver circadian clock reveals network dependency of metabolic rhythms
- PMID: 34550736
- PMCID: PMC8457671
- DOI: 10.1126/sciadv.abi7828
Integration of feeding behavior by the liver circadian clock reveals network dependency of metabolic rhythms
Abstract
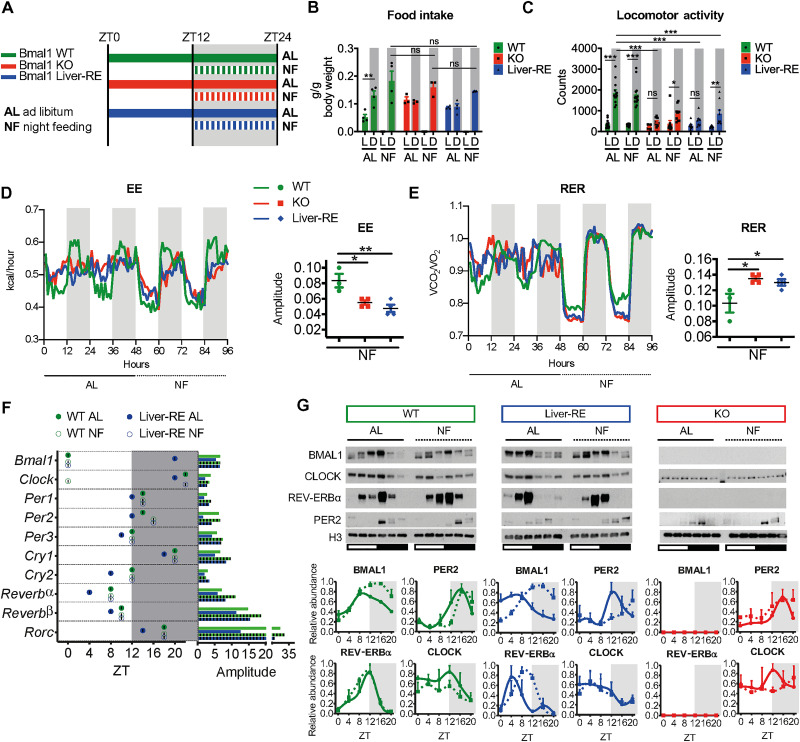
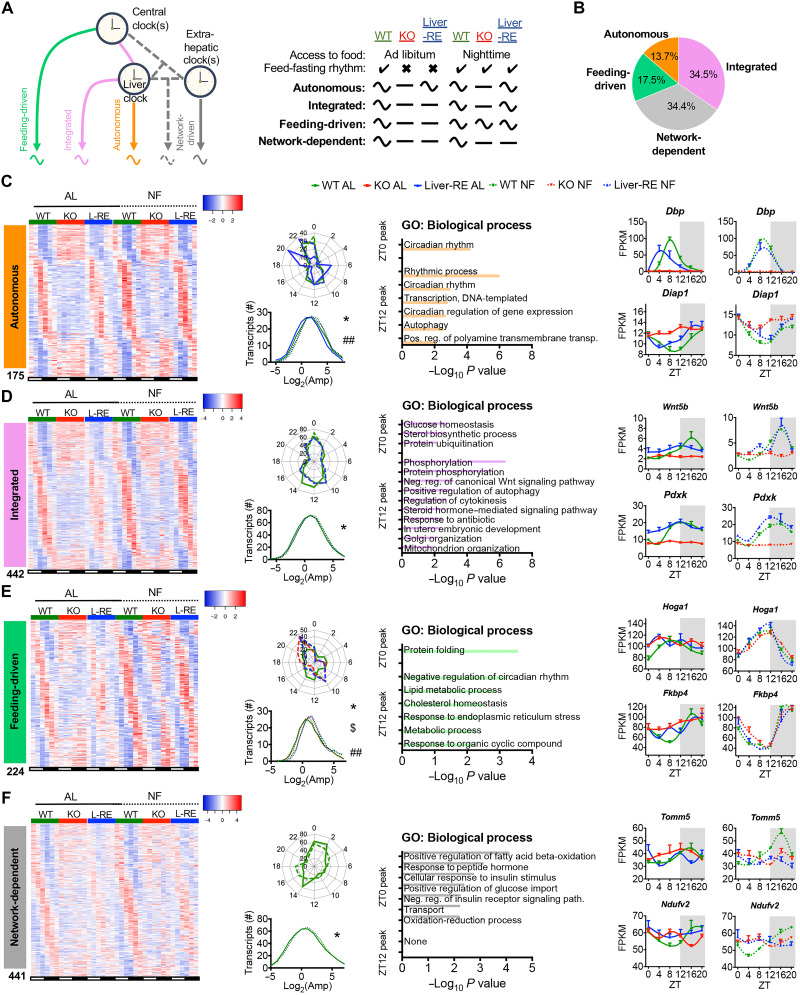
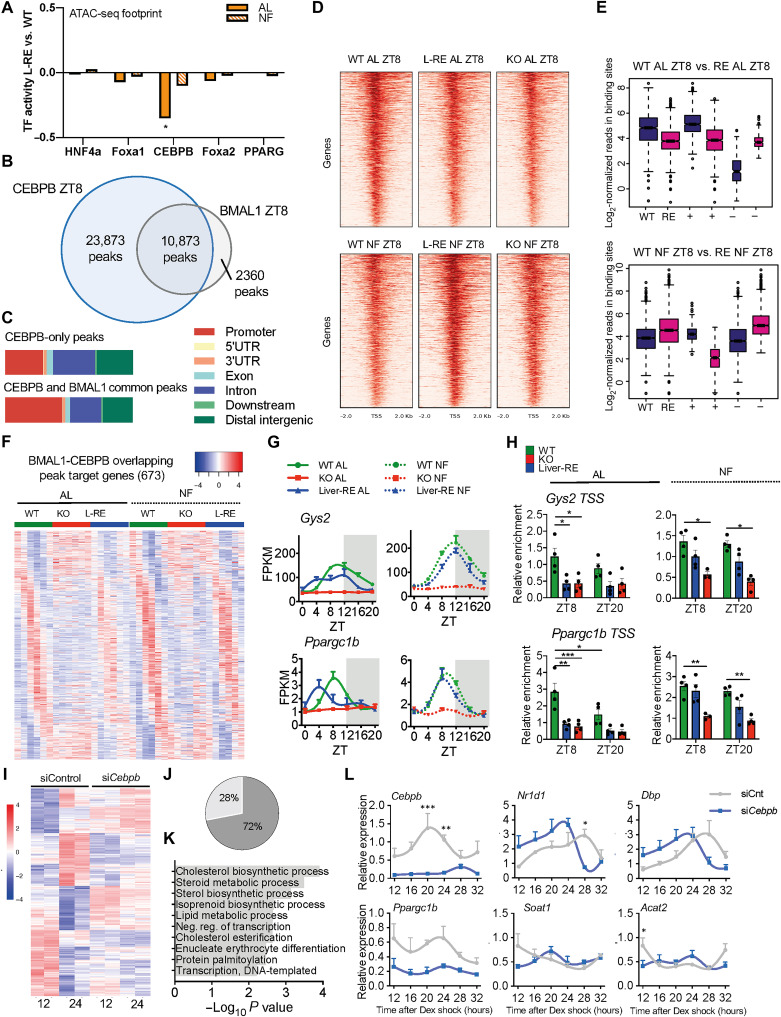
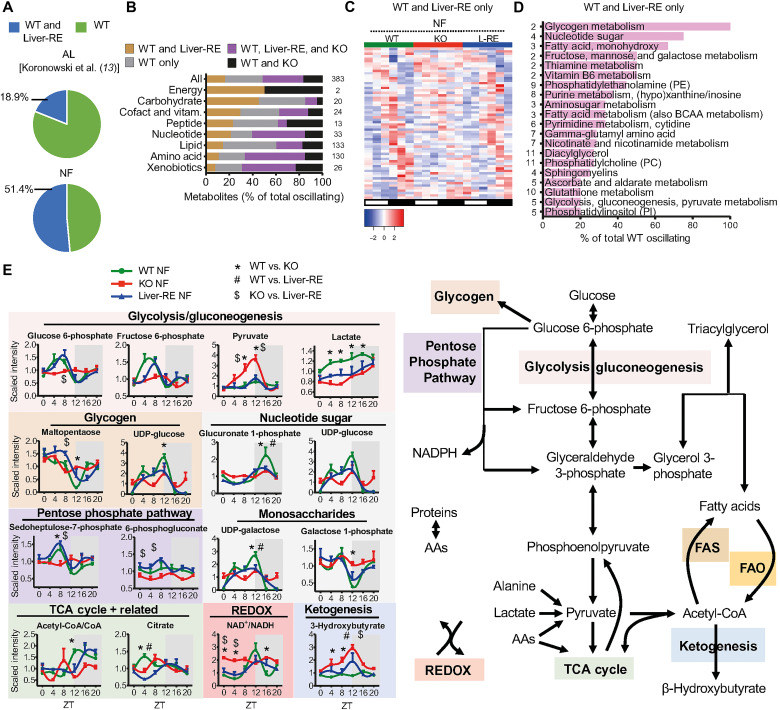
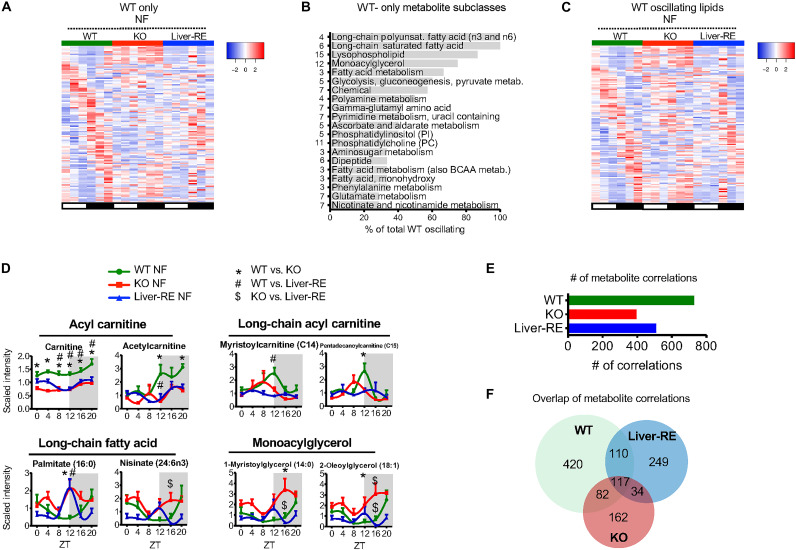
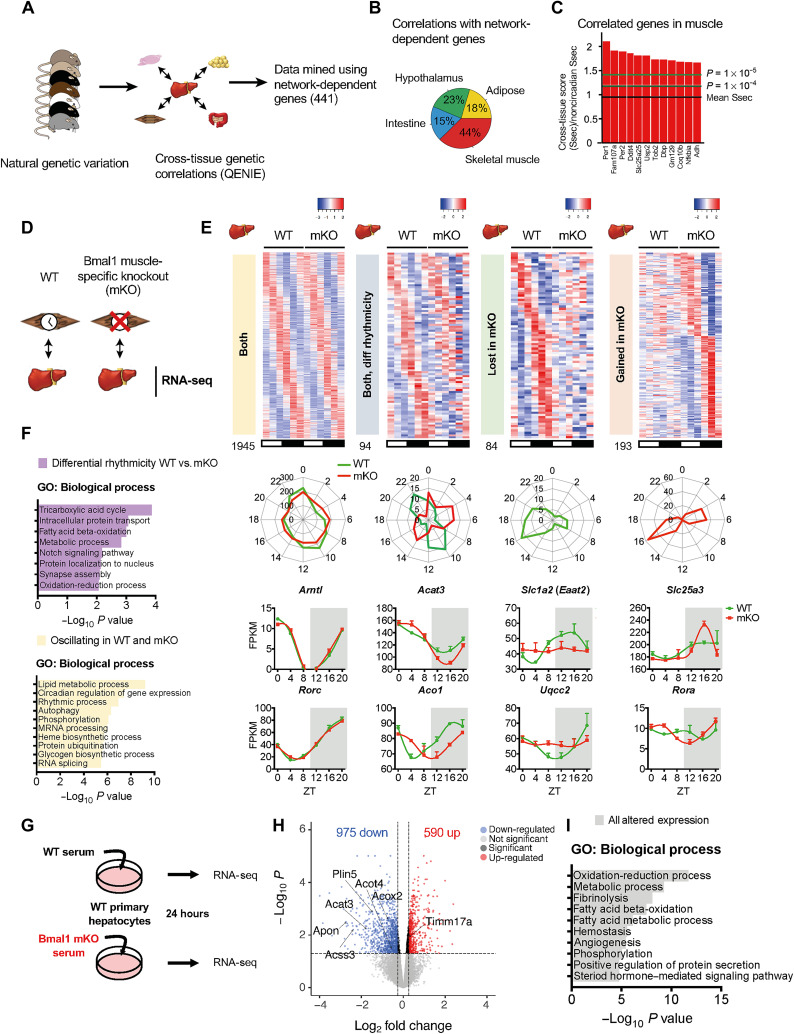
The mammalian circadian clock, expressed throughout the brain and body, controls daily metabolic homeostasis. Clock function in peripheral tissues is required, but not sufficient, for this task. Because of the lack of specialized animal models, it is unclear how tissue clocks interact with extrinsic signals to drive molecular oscillations. Here, we isolated the interaction between feeding and the liver clock by reconstituting Bmal1 exclusively in hepatocytes (Liver-RE), in otherwise clock-less mice, and controlling timing of food intake. We found that the cooperative action of BMAL1 and the transcription factor CEBPB regulates daily liver metabolic transcriptional programs. Functionally, the liver clock and feeding rhythm are sufficient to drive temporal carbohydrate homeostasis. By contrast, liver rhythms tied to redox and lipid metabolism required communication with the skeletal muscle clock, demonstrating peripheral clock cross-talk. Our results highlight how the inner workings of the clock system rely on communicating signals to maintain daily metabolism.
Figures
References
-
- Bass J., Lazar M. A., Circadian time signatures of fitness and disease. Science 354, 994–999 (2016). - PubMed
-
- Benitah S. A., Welz P. S., Circadian regulation of adult stem cell homeostasis and aging. Cell Stem Cell 26, 817–831 (2020). - PubMed
-
- Cederroth C. R., Albrecht U., Bass J., Brown S. A., Dyhrfjeld-Johnsen J., Gachon F., Green C. B., Hastings M. H., Helfrich-Förster C., Hogenesch J. B., Lévi F., Loudon A., Lundkvist G. B., Meijer J. H., Rosbash M., Takahashi J. S., Young M., Canlon B., Medicine in the fourth dimension. Cell Metab. 30, 238–250 (2019). - PMC - PubMed
Grants and funding
- R01 CA193466/CA/NCI NIH HHS/United States
- DP1 DK130640/DK/NIDDK NIH HHS/United States
- R00 HL138193/HL/NHLBI NIH HHS/United States
- T32 CA009054/CA/NCI NIH HHS/United States
- R01 CA228140/CA/NCI NIH HHS/United States
- S10 RR025496/RR/NCRR NIH HHS/United States
- R21 AG053592/AG/NIA NIH HHS/United States
- R21 DK114652/DK/NIDDK NIH HHS/United States
- S10 OD010794/OD/NIH HHS/United States
- P30 CA062203/CA/NCI NIH HHS/United States
- R01 HG007538/HG/NHGRI NIH HHS/United States
- K99 HL138193/HL/NHLBI NIH HHS/United States
- S10 OD021718/OD/NIH HHS/United States
- R01 GM123558/GM/NIGMS NIH HHS/United States
- F32 DK121425/DK/NIDDK NIH HHS/United States
