

Deletion of pleiotrophin impairs glucose tolerance and liver metabolism in pregnant mice: Moonlighting role of glycerol kinase
- PMID: 34551152
- PMCID: PMC12316093
- DOI: 10.1096/fj.202101181R
Deletion of pleiotrophin impairs glucose tolerance and liver metabolism in pregnant mice: Moonlighting role of glycerol kinase
Abstract
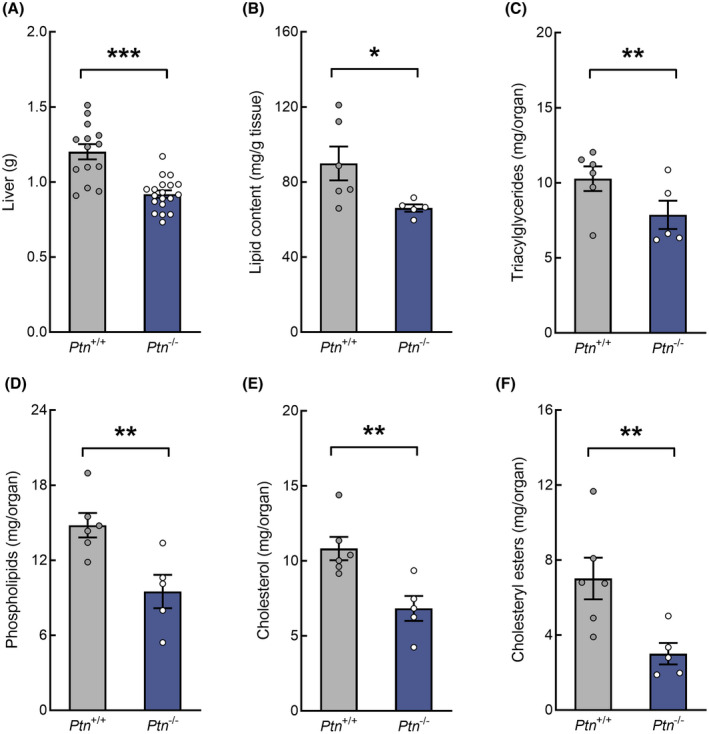
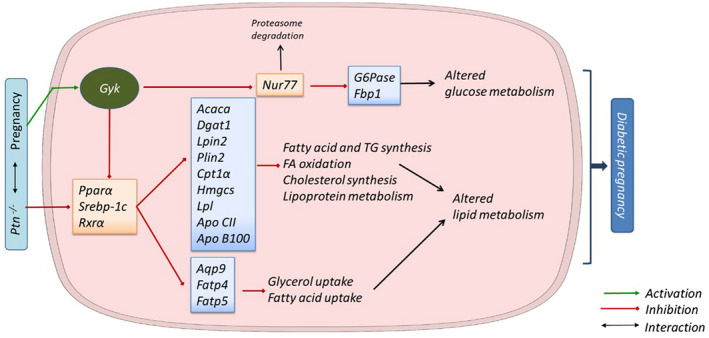
Pleiotrophin is a pleiotropic cytokine that has been demonstrated to have a critical role in regulating energy metabolism, lipid turnover and plasticity of adipose tissue. Here, we hypothesize that this cytokine can be involved in regulatory processes of glucose and lipid homeostasis in the liver during pregnancy. Using 18-days pregnant Ptn-deficient mice, we evaluated the biochemical profile (circulating variables), tissue mRNA expression (qPCR) and protein levels of key enzymes and transcription factors involved in main metabolic pathways. Ptn deletion was associated with a reduction in body weight gain, hyperglycemia and glucose intolerance. Moreover, we observed an impairment in glucose synthesis and degradation during late pregnancy in Ptn-/- mice. Hepatic lipid content was significantly lower (73.6%) in Ptn-/- mice and was associated with a clear reduction in fatty acid, triacylglycerides and cholesterol synthesis. Ptn deletion was accompanying with a diabetogenic state in the mother and a decreased expression of key proteins involved in glucose and lipid uptake and metabolism. Moreover, Ptn-/- pregnant mice have a decreased expression of transcription factors, such as PPAR-α, regulating lipid uptake and glucose and lipid utilization. Furthermore, the augmented expression and nuclear translocation of glycerol kinase, and the decrease in NUR77 protein levels in the knock-out animals can further explain the alterations observed in hepatic glucose metabolism. Our results point out for the first time that pleiotrophin is an important player in maintaining hepatic metabolic homeostasis during late gestation, and further highlighted the moonlighting role of glycerol kinase in the regulation of maternal glucose homeostasis during pregnancy.
Keywords: NR4A1; glycerol kinase; liver; metabolism; pleiotrophin; pregnancy.
© 2021 The Authors. The FASEB Journal published by Wiley Periodicals LLC on behalf of Federation of American Societies for Experimental Biology.
Conflict of interest statement
The authors declared that no conflicts of interest exist with this manuscript.
Figures







References
-
- Herrera E. Metabolic adaptations in pregnancy and their implications for the availability of substrates to the fetus. Eur J Clin Nutr. 2000;54(Suppl 1):S47‐S51. - PubMed
-
- Ramos MP, Crespo‐Solans MD, del Campo S, Cacho J, Herrera E. Fat accumulation in the rat during early pregnancy is modulated by enhanced insulin responsiveness. Am J Physiol Endocrinol Metab. 2003;285:E318‐E328. - PubMed
-
- Herrera E, del Campo S, Marciniak J, Sevillano J, Ramos MP. Enhanced utilization of glycerol for glyceride synthesis in isolated adipocytes from early pregnant rats. J Physiol Biochem. 2010;66:245‐253. - PubMed
-
- Villar J, Cogswell M, Kestler E, Castillo P, Menendez R, Repke JT. Effect of fat and fat‐free mass deposition during pregnancy on birth weight. Am J Obstet Gynecol. 1992;167:1344‐1352. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
