

Review
doi: 10.1038/s41589-021-00807-5.
Epub 2021 Sep 22.
Escherichia coli small molecule metabolism at the host-microorganism interface
Affiliations
- PMID: 34552219
- PMCID: PMC8675634
- DOI: 10.1038/s41589-021-00807-5
Item in Clipboard
Review
Escherichia coli small molecule metabolism at the host-microorganism interface
Nat Chem Biol.
2021 Oct.
Abstract
Escherichia coli are a common component of the human microbiota, and isolates exhibit probiotic, commensal and pathogenic roles in the host. E. coli members often use diverse small molecule chemistry to regulate intrabacterial, intermicrobial and host-bacterial interactions. While E. coli are considered to be a well-studied model organism in biology, much of their chemical arsenal has only more recently been defined, and much remains to be explored. Here we describe chemical signaling systems in E. coli in the context of the broader field of metabolism at the host-bacteria interface and the role of this signaling in disease modulation.
© 2021. Springer Nature America, Inc.
Figures
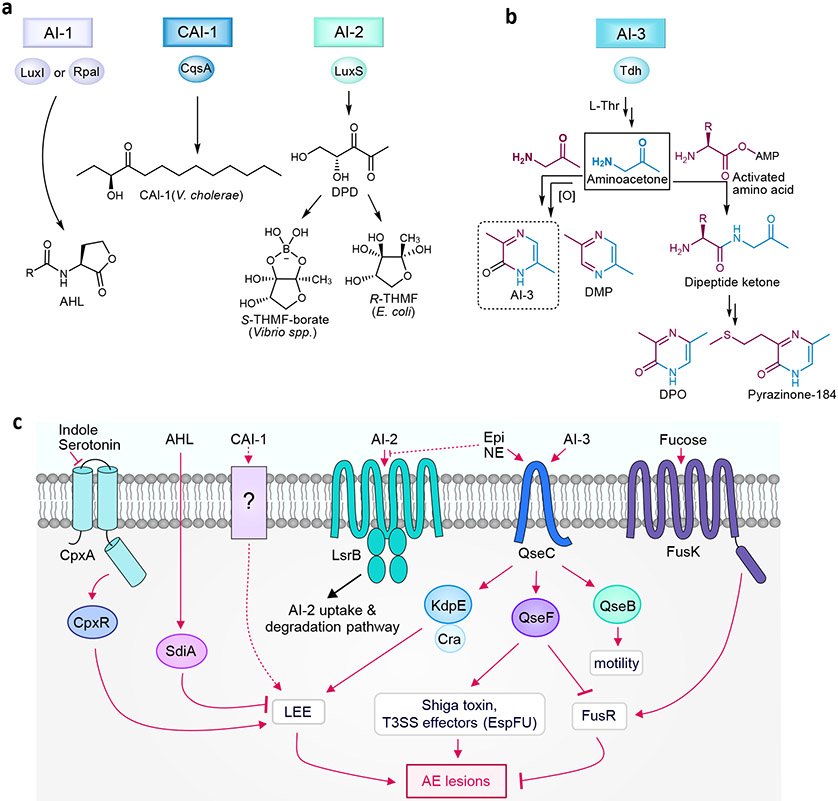
a∣ Synthases and structures of autoinducer-1 (N-acyl homoserine lactone (AHL)), Vibrio cholerae autoinducer-1 (CAI-1), and autoinducer-2 (AI-2). b∣ Biosynthesis of autoinducer-3 (AI-3), 2,5-dimethylpyrazine (DMP), 3,5-dimethyl-pyrazin-2-one (DPO), and analogs. c∣ Quorum sensing receptors and signaling pathways in E. coli. Epi, epinephrine. NE, norepinephrine.
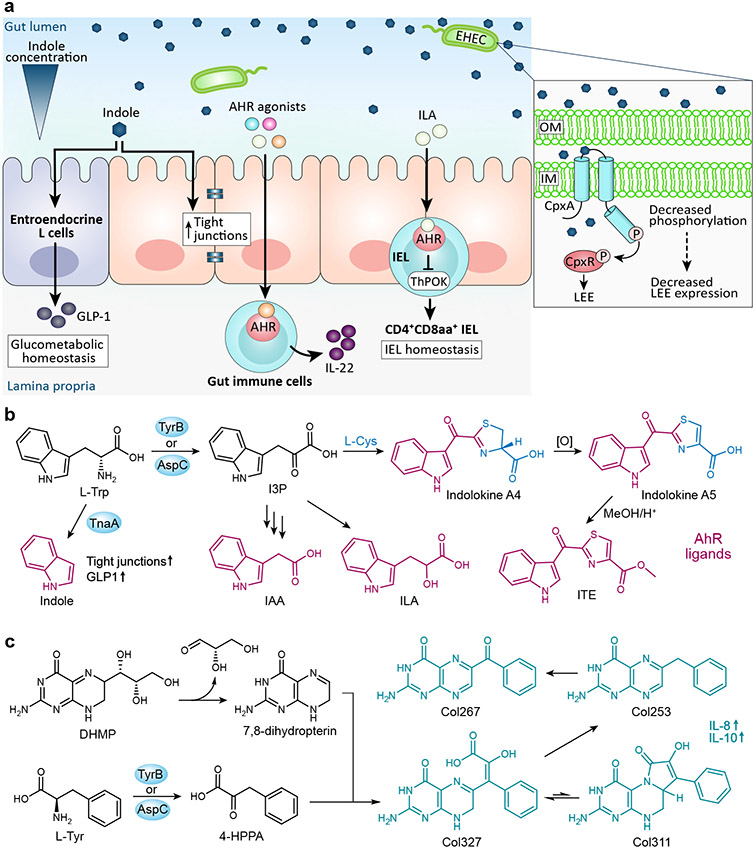
a∣ Here we highlight a few of the signaling pathways induced by E. coli tryptophan catabolites. Indole stimulates tight junctions and glucagon-like peptide 1 (GLP-1), respectively, affecting intestinal barrier integrity and glycometabolism homeostasis. An indole concentration gradient in the human gut influences the virulence gene expression in EHEC, with the expression of the LEE pathogenicity island decreasing in the presence of indole. Several tryptophan catabolites act as aryl hydrocarbon receptor (AhR) ligands to influence host immune response. Indole-3-lactic acid (ILA) stimulates the development of CD4+CD8αα+ intraepithelial lymphocytes. b∣ Tryptophan catabolites produced by E. coli that also serve as AhR ligands include indole, indole acetic acid (IAA), and ILA. E. coli also produce the AhR ligands indolokine A4 and indolokine A5, which are formed from indole-3-pyruvic acid (I3P) and L-cysteine, and resemble the potent AhR agonist 2-(1′H-indole-3′-carbonyl)-thiazole-4-carboxylic acid methyl ester (ITE). c∣ The tryptophan derivative 3-(4-hydroxyphenyl)pyruvate (4-HPPA) reacts with 7,8 dihydropterin, a degradation product of the folate/monapterin pathway, to form a molecular family termed the colipterins.
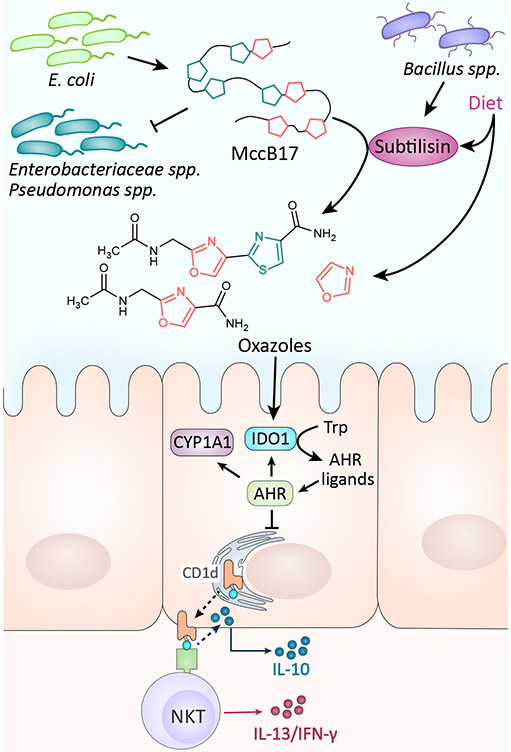
MccB17 is produced by E. coli strains to inhibit the growth of closely related bacteria by targeting their DNA gyrase. MccB17 also can be cleaved by the protease subtilisin to form small oxazole containing fragments. Oxazoles derived from diet and microbes activate indoleamine 2,3-dioxygenase (IDO1) which induces the production of AhR ligands in intestinal epithelial cells (IECs). AhR activation in IECs leads to invariant natural killer T (iNKT) cell-mediated intestinal inflammation.
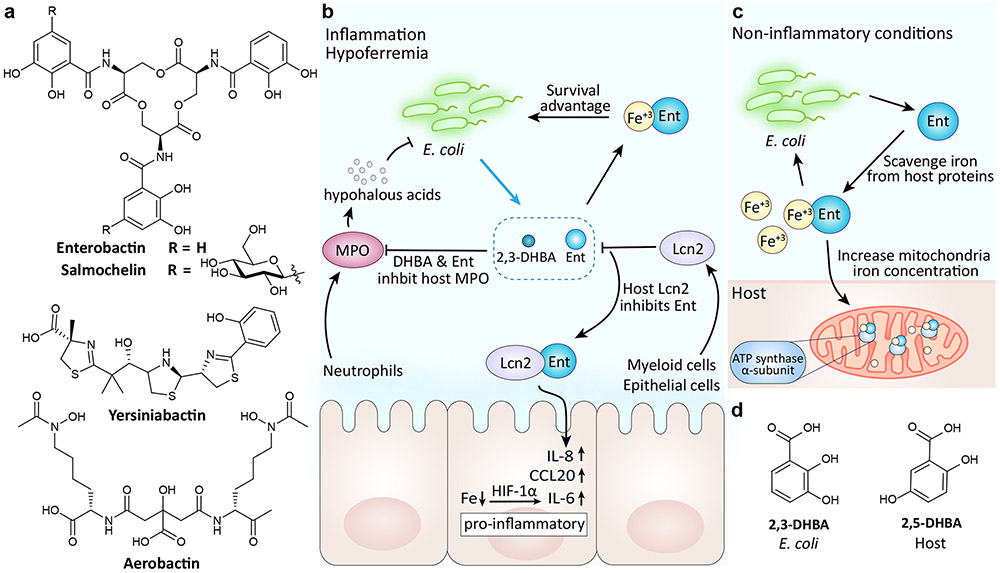
a∣ Structures of E. coli siderophores. b∣ During inflammation, environmental iron levels decrease. In turn, enterobactin secretion is increased to maintain E. coli’s iron load. Enterobactin and its precursor 2,3-dihydroxybenzoic acid (DHBA) also promote E. coli survival by inhibiting the bactericidal enzyme myeloperoxidase (MPO). As a countermeasure, the host protein lipocalin 2 (Lcn2) binds enterobactin. The Lcn2-enterobactin complex prevents the pirating of iron and induces a host immune response. c∣ As part of a mutualistic relationship between healthy gut bacteria and the host, enterobactin promotes mitochondrial iron uptake by binding the to host ATP synthase α subunit. d∣ Structure of bacterial 2,3-DHBA and its human analog 2,5-DHBA.
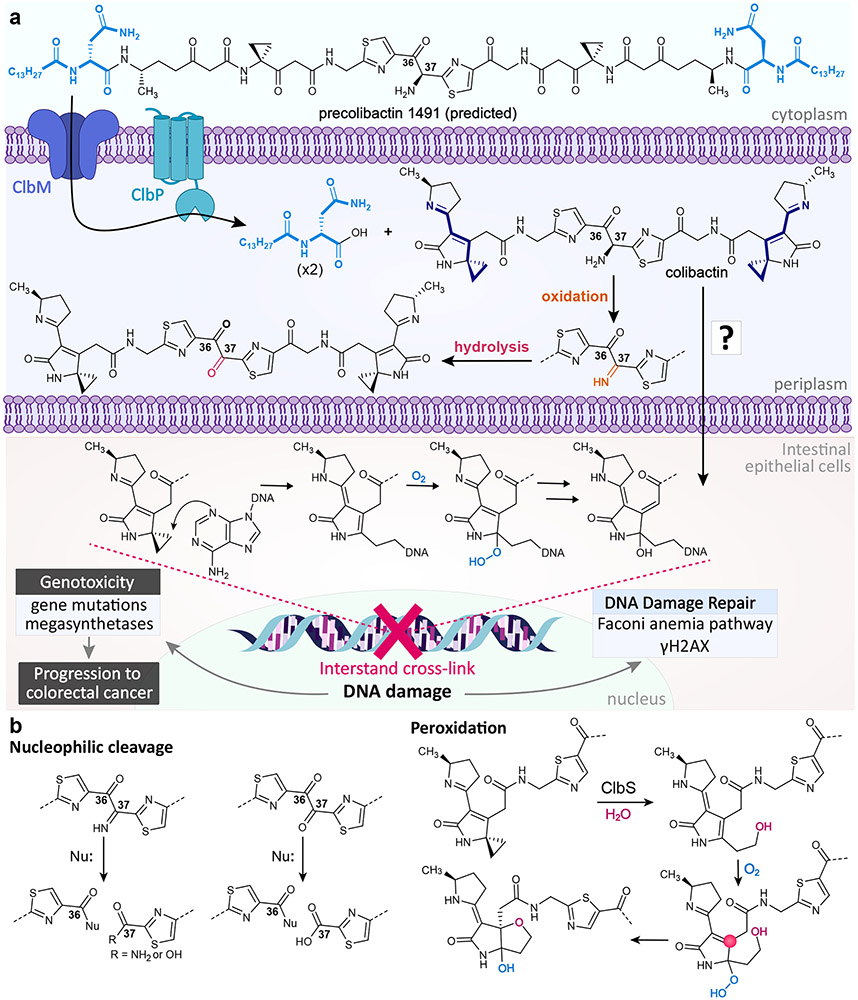
a∣ Precolibactin 1491 is transported into the periplasm and cleaved to form a mature colibactin. The α-amino-ketone bithiazole spacer is subject to autoxidation at the C37 position in vitro, though it is unclear to what extent oxidation would occur in vivo. Colibactin is transported into host epithelial cells through an unknown mechanism, where its electrophilic spirocyclopropane moieties cross-link DNA. b∣ In the absence of ClbP, precolibactin 1491 undergoes oxidation and macrocyclization to form precolibactin 1489, a prediction based on tandem MS, isotopic labeling, genome editing studies, and smaller characterized macrocycles. c∣ Colibactin has been observed to undergo reactions other than DNA cross-linking, such as nucleophilic cleavage of the oxidized α-amino-ketone bithiazole spacer and peroxidation. In the peroxidation mechanism, the C4 position (pink circle) can act as an electrophilic site, although it can also undergo stable tautomerization.
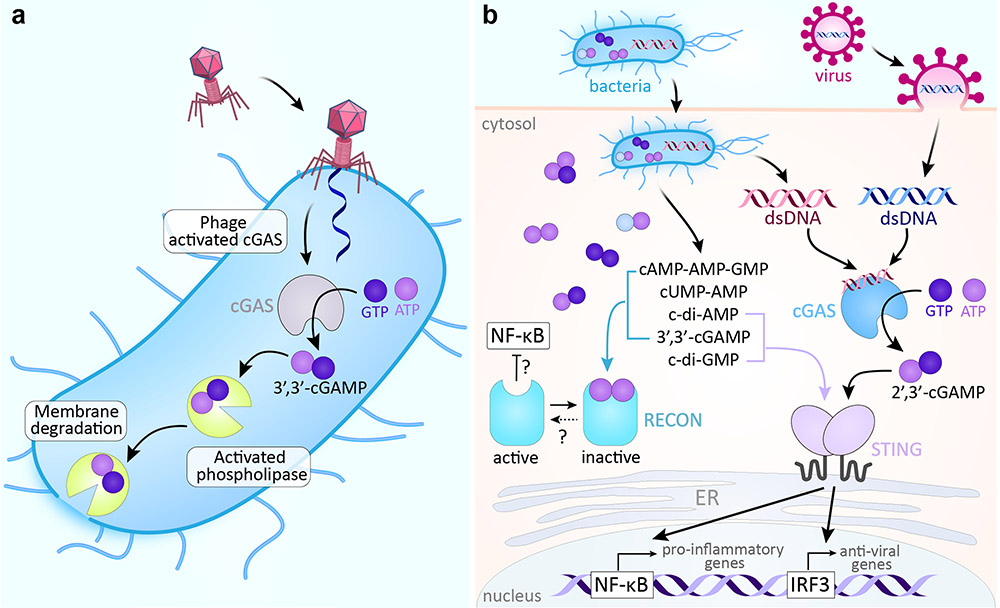
a∣ In CBASS, phage infection triggers the synthesis of 3’,3’-cGAMP, which then activates phospholipase. The phospholipase degrades the bacterial membrane, leading to altruistic suicide prior to the completion of phage reproduction and thus preventing the further spread of infection. b∣ In the mammalian cGAS–STING pathway, cGAS senses the DNA of bacterial and viral invaders and synthesizes 2’,3’-cGAMP, which in turn binds and activates STING. STING activates transcription factors needed for downstream transcription of innate immune response genes. The bacterial cdNs, c-di-GMP and 3’,3’-cGAMP, also directly activate STING. Several bacterial signals bind to RECON, inhibiting its undefined oxidoreductase activity and leading to increased activation of the proinflammatory transcription factor NF-κB. CBASS, cyclic-oligonucleotide-based anti-phage signaling system; cGAMP, cyclic GMP-AMP; cGAS, cyclic GMP-AMP synthase; STING, stimulator of interferon genes; cdN, cyclic dinucletotide; RECON, reductase controlling NF-κB.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
