

Potent anti-tumor effects of receptor-retargeted syncytial oncolytic herpes simplex virus
- PMID: 34553018
- PMCID: PMC8426171
- DOI: 10.1016/j.omto.2021.08.002
Potent anti-tumor effects of receptor-retargeted syncytial oncolytic herpes simplex virus
Abstract
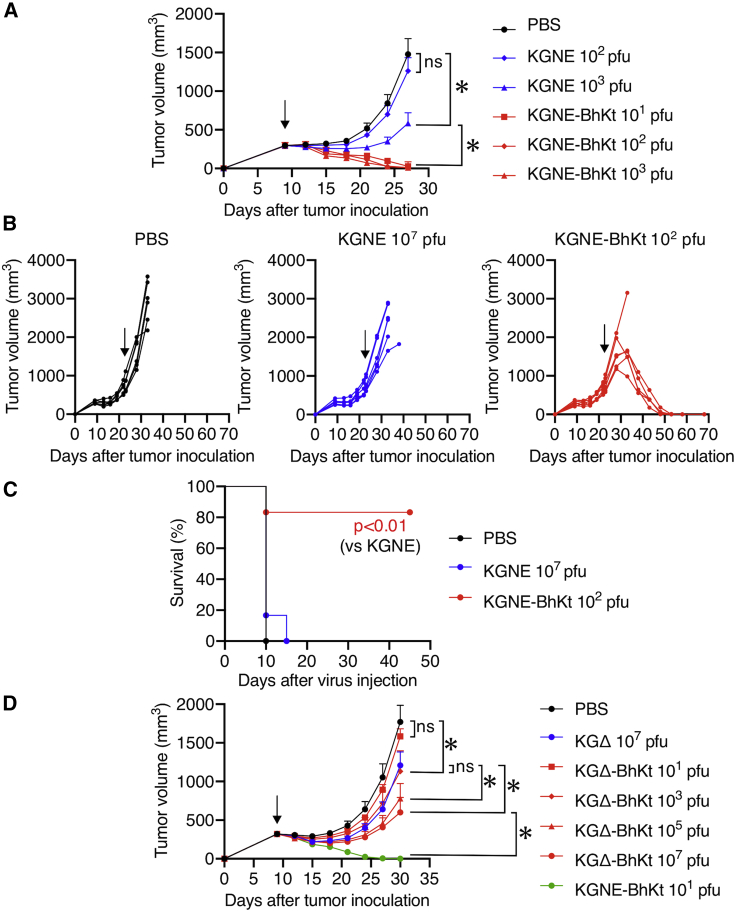
Most oncolytic virotherapy has thus far employed viruses deficient in genes essential for replication in normal cells but not in cancer cells. Intra-tumoral injection of such viruses has resulted in clinically significant anti-tumor effects on the lesions in the vicinity of the injection sites but not on distant visceral metastases. To overcome this limitation, we have developed a receptor-retargeted oncolytic herpes simplex virus employing a single-chain antibody for targeting tumor-associated antigens (RR-oHSV) and its modified version with additional mutations conferring syncytium formation (RRsyn-oHSV). We previously showed that RRsyn-oHSV exhibits preserved antigen specificity and an ∼20-fold higher tumoricidal potency in vitro relative to RR-oHSV. Here, we investigated the in vivo anti-tumor effects of RRsyn-oHSV using human cancer xenografts in immunodeficient mice. With only a single intra-tumoral injection of RRsyn-oHSV at very low doses, all treated tumors regressed completely. Furthermore, intra-venous administration of RRsyn-oHSV resulted in robust anti-tumor effects even against large tumors. We found that these potent anti-tumor effects of RRsyn-oHSV may be associated with the formation of long-lasting tumor cell syncytia not containing non-cancerous cells that appear to trigger death of the syncytia. These results strongly suggest that cancer patients with distant metastases could be effectively treated with our RRsyn-oHSV.
Keywords: HSV; cancer; cell death; fusogenic; gene therapy; herpes simplex virus; intravenous; oncolytic virotherapy; systemic delivery; targeting.
© 2021 The Author(s).
Conflict of interest statement
H.U. is an inventor of intellectual property licensed to Oncorus, Inc. (Cambridge, MA, USA). R.H. and K.O. are employees of Ono Pharmaceutical Co., Ltd.
Figures







Similar articles
-
Induction of necroptosis in multinucleated giant cells induced by conditionally replicating syncytial oHSV in co-cultures of cancer cells and non-cancerous cells.Mol Ther Oncol. 2024 Apr 15;32(2):200803. doi: 10.1016/j.omton.2024.200803. eCollection 2024 Jun 20. Mol Ther Oncol. 2024. PMID: 38706990 Free PMC article.
-
Systemic delivery of glycosylated-PEG-masked oncolytic virus enhances targeting of antitumor immuno-virotherapy and modulates T and NK cell infiltration.Theranostics. 2023 Oct 2;13(15):5452-5468. doi: 10.7150/thno.87498. eCollection 2023. Theranostics. 2023. PMID: 37908722 Free PMC article.
-
Syncytial Mutations Do Not Impair the Specificity of Entry and Spread of a Glycoprotein D Receptor-Retargeted Herpes Simplex Virus.J Virol. 2016 Nov 28;90(24):11096-11105. doi: 10.1128/JVI.01456-16. Print 2016 Dec 15. J Virol. 2016. PMID: 27707922 Free PMC article.
-
Oncolytic herpes simplex virus and immunotherapy.BMC Immunol. 2018 Dec 18;19(1):40. doi: 10.1186/s12865-018-0281-9. BMC Immunol. 2018. PMID: 30563466 Free PMC article. Review.
-
Oncolytic herpes simplex virus-based strategies: toward a breakthrough in glioblastoma therapy.Front Microbiol. 2014 Jun 20;5:303. doi: 10.3389/fmicb.2014.00303. eCollection 2014. Front Microbiol. 2014. PMID: 24999342 Free PMC article. Review.
Cited by
-
Oncolytic herpes simplex viruses designed for targeted treatment of EGFR-bearing tumors.Mol Ther Oncol. 2024 Jan 10;32(1):200761. doi: 10.1016/j.omton.2024.200761. eCollection 2024 Mar 21. Mol Ther Oncol. 2024. PMID: 38596286 Free PMC article.
-
Replication and Spread of Oncolytic Herpes Simplex Virus in Solid Tumors.Viruses. 2022 Jan 10;14(1):118. doi: 10.3390/v14010118. Viruses. 2022. PMID: 35062322 Free PMC article. Review.
-
HSV: The scout and assault for digestive system tumors.Front Mol Biosci. 2023 Feb 28;10:1142498. doi: 10.3389/fmolb.2023.1142498. eCollection 2023. Front Mol Biosci. 2023. PMID: 36926680 Free PMC article. Review.
-
Comparison of the oncolytic activity of a replication-competent and a replication-deficient herpes simplex virus 1.Immunology. 2024 Jun;172(2):279-294. doi: 10.1111/imm.13775. Epub 2024 Mar 5. Immunology. 2024. PMID: 38444199 Free PMC article.
-
Immune landscape and response to oncolytic virus-based immunotherapy.Front Med. 2024 Jun;18(3):411-429. doi: 10.1007/s11684-023-1048-0. Epub 2024 Mar 8. Front Med. 2024. PMID: 38453818 Review.
References
-
- Rampling R., Cruickshank G., Papanastassiou V., Nicoll J., Hadley D., Brennan D., Petty R., MacLean A., Harland J., McKie E. Toxicity evaluation of replication-competent herpes simplex virus (ICP 34.5 null mutant 1716) in patients with recurrent malignant glioma. Gene Ther. 2000;7:859–866. - PubMed
-
- Markert J.M., Medlock M.D., Rabkin S.D., Gillespie G.Y., Todo T., Hunter W.D., Palmer C.A., Feigenbaum F., Tornatore C., Tufaro F., Martuza R.L. Conditionally replicating herpes simplex virus mutant, G207 for the treatment of malignant glioma: results of a phase I trial. Gene Ther. 2000;7:867–874. - PubMed
-
- Andtbacka R.H., Kaufman H.L., Collichio F., Amatruda T., Senzer N., Chesney J., Delman K.A., Spitler L.E., Puzanov I., Agarwala S.S. Talimogene Laherparepvec Improves Durable Response Rate in Patients With Advanced Melanoma. J. Clin. Oncol. 2015;33:2780–2788. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
