

Characterization of changes in the hemagglutinin that accompanied the emergence of H3N2/1968 pandemic influenza viruses
- PMID: 34555124
- PMCID: PMC8491938
- DOI: 10.1371/journal.ppat.1009566
Characterization of changes in the hemagglutinin that accompanied the emergence of H3N2/1968 pandemic influenza viruses
Abstract
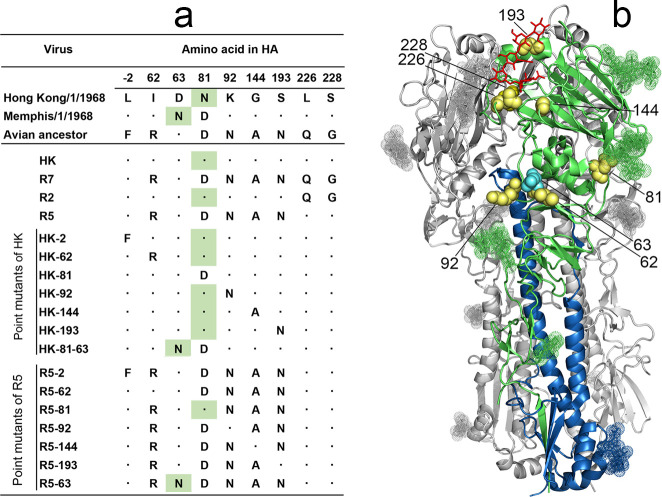
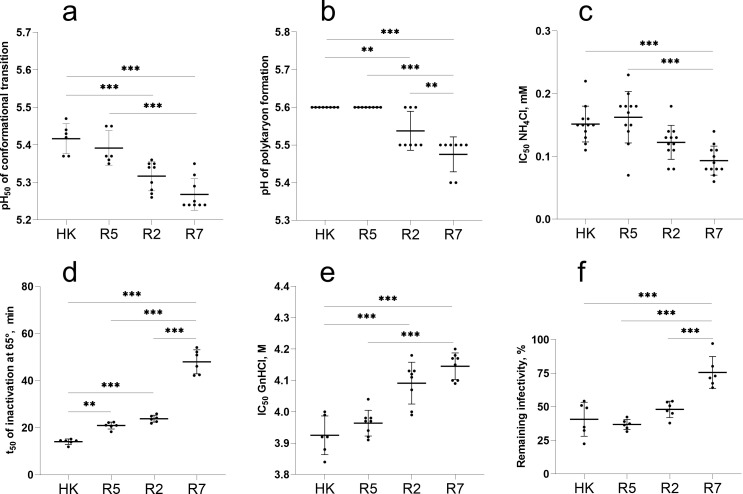
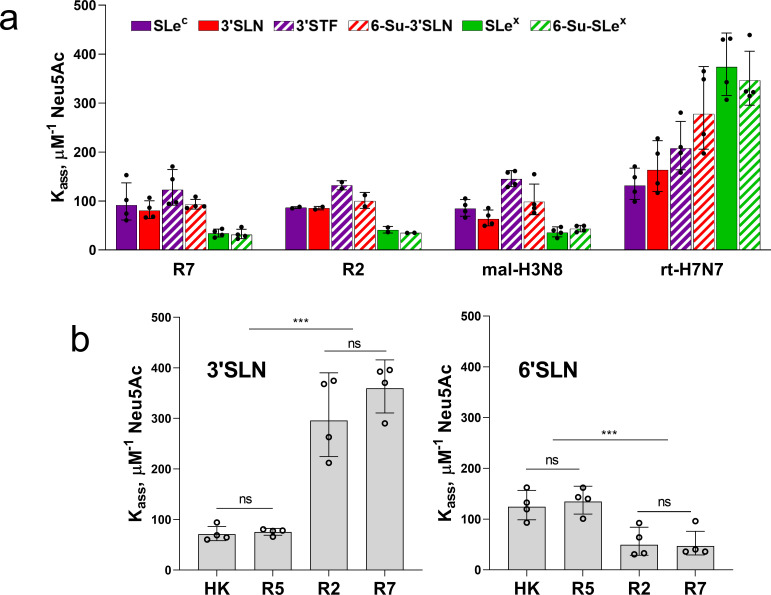
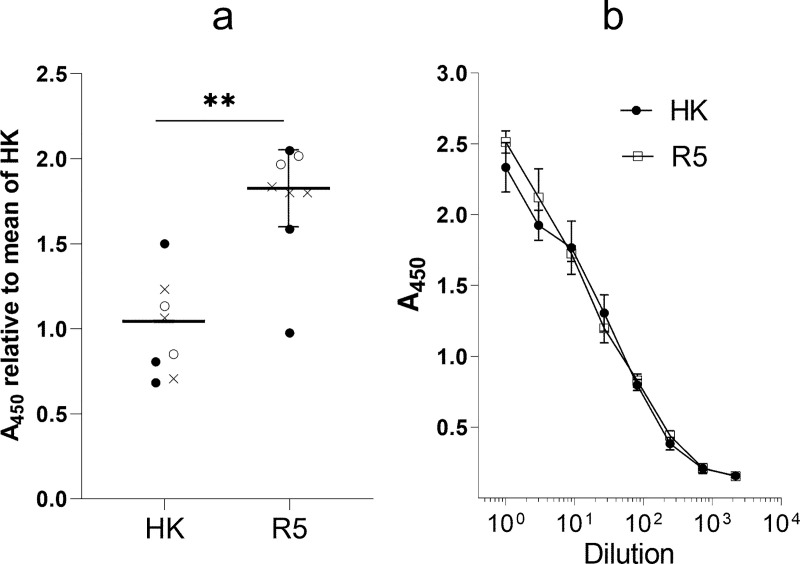
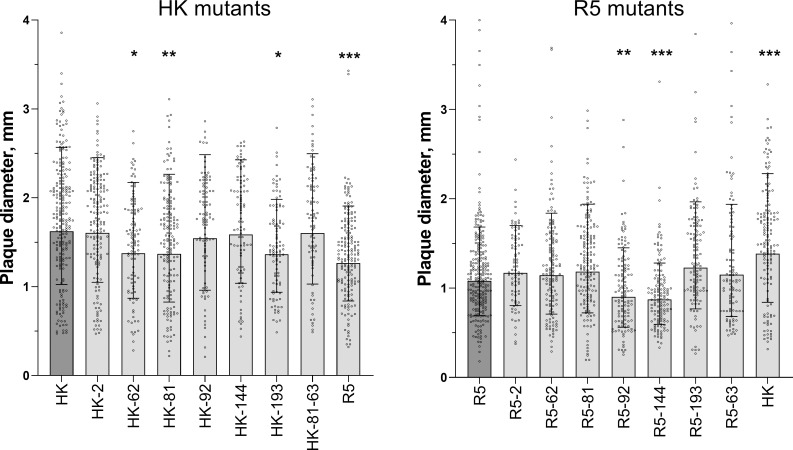
The hemagglutinin (HA) of A/H3N2 pandemic influenza viruses (IAVs) of 1968 differed from its inferred avian precursor by eight amino acid substitutions. To determine their phenotypic effects, we studied recombinant variants of A/Hong Kong/1/1968 virus containing either human-type or avian-type amino acids in the corresponding positions of HA. The precursor HA displayed receptor binding profile and high conformational stability typical for duck IAVs. Substitutions Q226L and G228S, in addition to their known effects on receptor specificity and replication, marginally decreased HA stability. Substitutions R62I, D63N, D81N and N193S reduced HA binding avidity. Substitutions R62I, D81N and A144G promoted viral replication in human airway epithelial cultures. Analysis of HA sequences revealed that substitutions D63N and D81N accompanied by the addition of N-glycans represent common markers of avian H3 HA adaptation to mammals. Our results advance understanding of genotypic and phenotypic changes in IAV HA required for avian-to-human adaptation and pandemic emergence.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
