

Follicular dendritic cells restrict interleukin-4 availability in germinal centers and foster memory B cell generation
- PMID: 34555336
- PMCID: PMC8516727
- DOI: 10.1016/j.immuni.2021.08.028
Follicular dendritic cells restrict interleukin-4 availability in germinal centers and foster memory B cell generation
Abstract
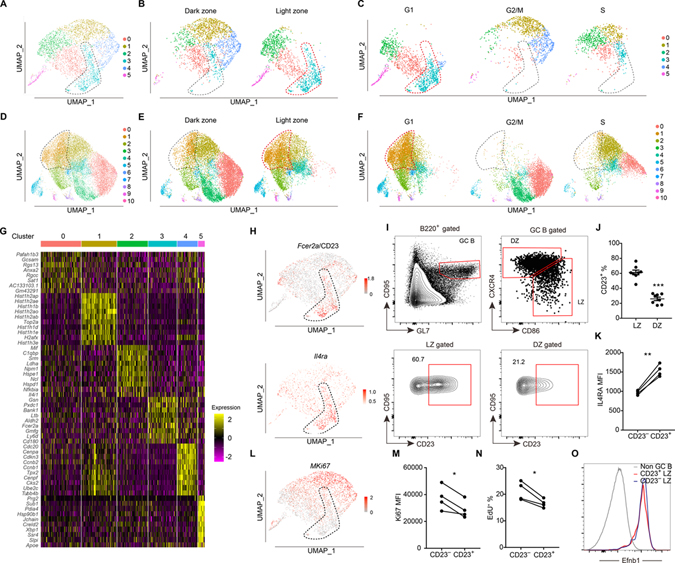
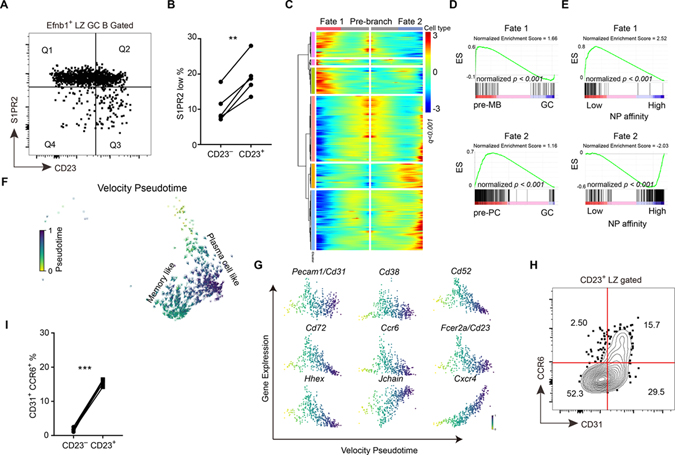
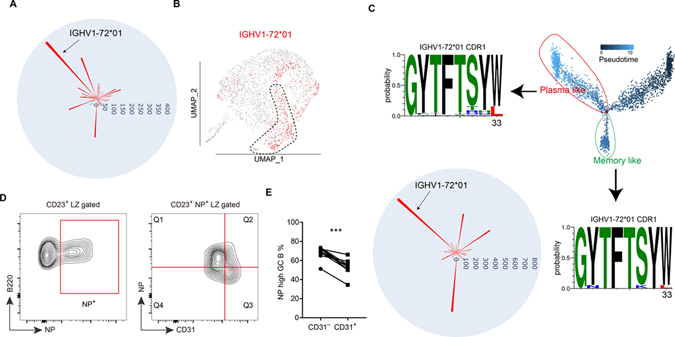
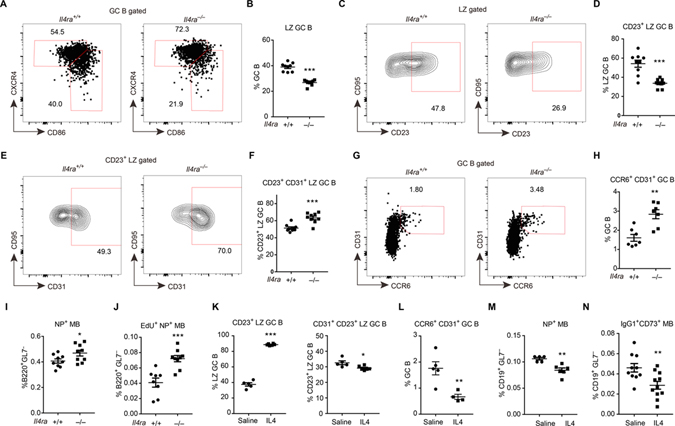
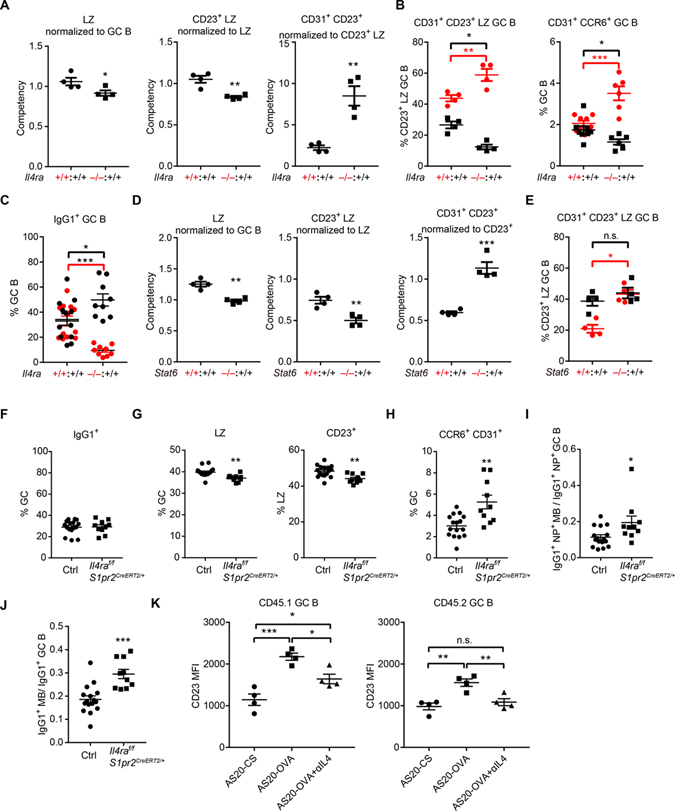
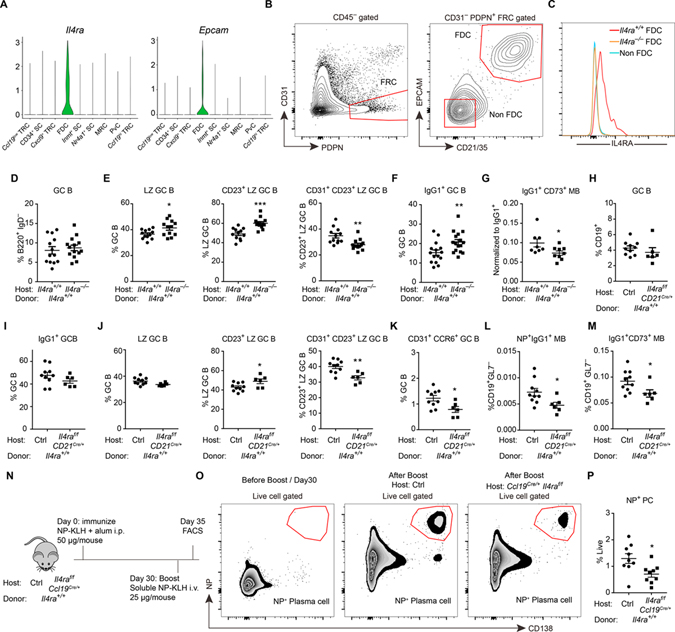
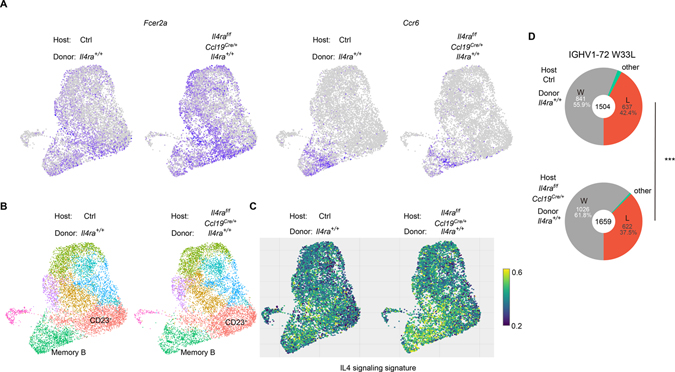
B cells within germinal centers (GCs) enter cycles of antibody affinity maturation or exit the GC as memory cells or plasma cells. Here, we examined the contribution of interleukin (IL)-4 on B cell fate decisions in the GC. Single-cell RNA-sequencing identified a subset of light zone GC B cells expressing high IL-4 receptor-a (IL4Ra) and CD23 and lacking a Myc-associated signature. These cells could differentiate into pre-memory cells. B cell-specific deletion of IL4Ra or STAT6 favored the pre-memory cell trajectory, and provision of exogenous IL-4 in a wild-type context reduced pre-memory cell frequencies. IL-4 acted during antigen-specific interactions but also influenced bystander cells. Deletion of IL4Ra from follicular dendritic cells (FDCs) increased the availability of IL-4 in the GC, impaired the selection of affinity-matured B cells, and reduced memory cell generation. We propose that GC FDCs establish a niche that limits bystander IL-4 activity, focusing IL-4 action on B cells undergoing selection and enhancing memory cell differentiation.
Keywords: IL-4; affinity maturation; follicular dendritic cells; germinal center; memory B cells; scRNA-seq; scVDJ-seq.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.G.C. is an Scientific Advisory Board member of Be Biopharma and MiroBio.
Figures
References
-
- Andoh A, Masuda A, Yamakawa M, Kumazawa Y, and Kasajima T (2000). Absence of interleukin-4 enhances germinal center reaction in secondary immune response. Immunol. Lett. 73, 35–41. - PubMed
-
- Bannard O, and Cyster JG (2017). Germinal centers: programmed for affinity maturation and antibody diversification. Curr. Opin. Immunol. 45, 21–30. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
