

Role of pyruvate in maintaining cell viability and energy production under high-glucose conditions
- PMID: 34556698
- PMCID: PMC8460646
- DOI: 10.1038/s41598-021-98082-w
Role of pyruvate in maintaining cell viability and energy production under high-glucose conditions
Abstract
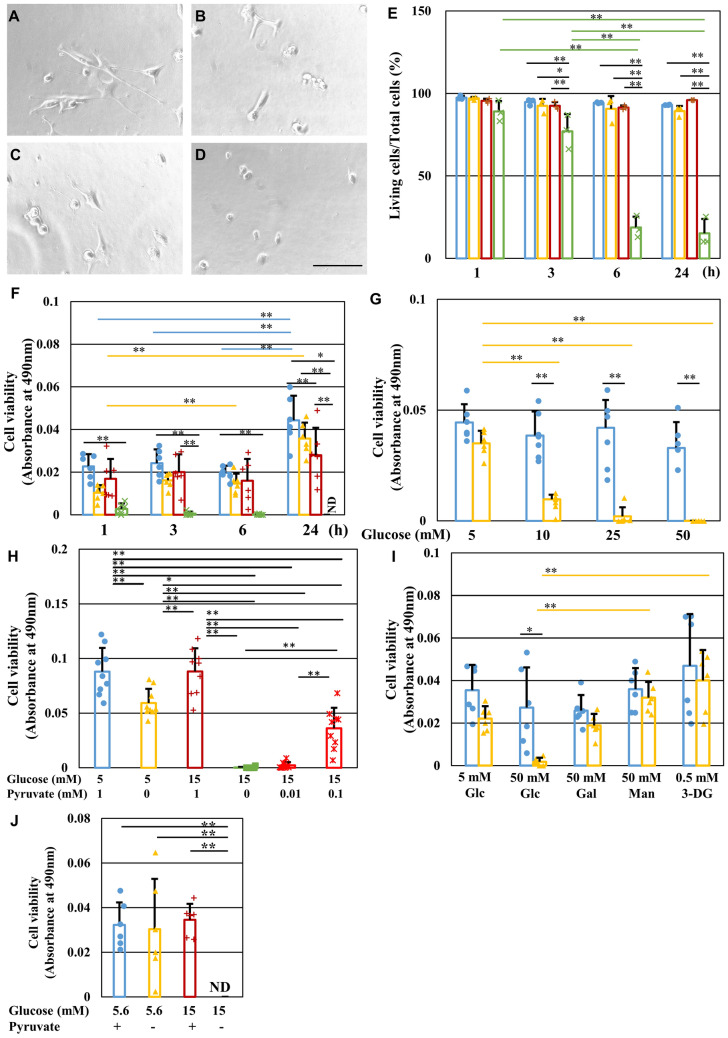
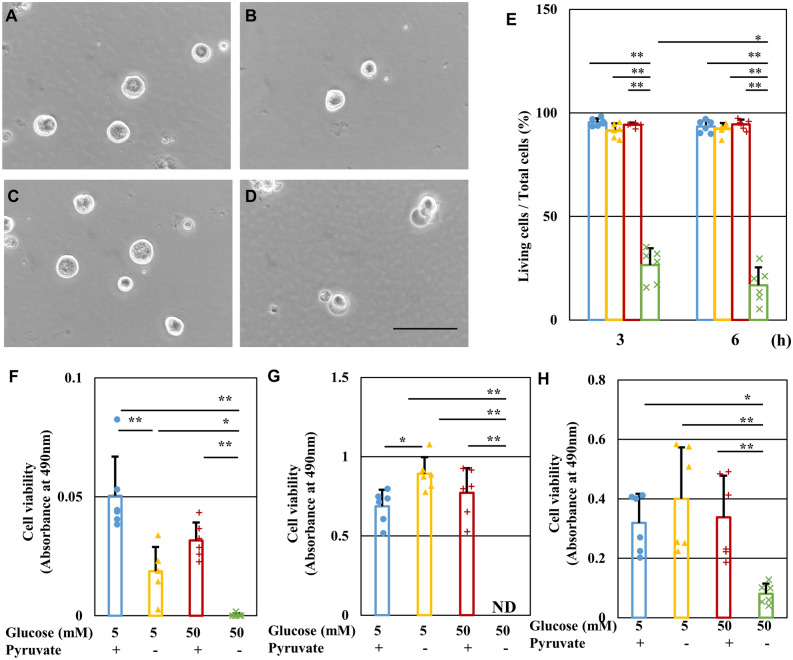
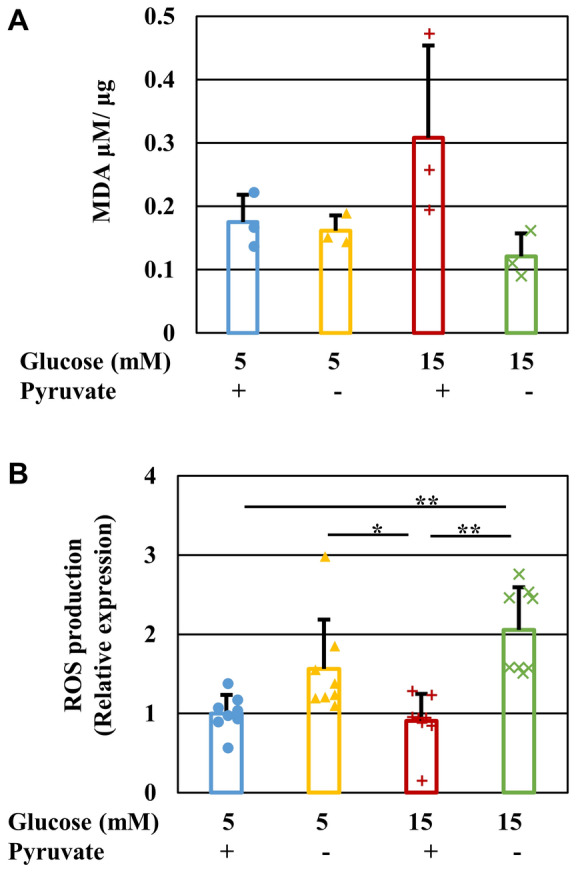
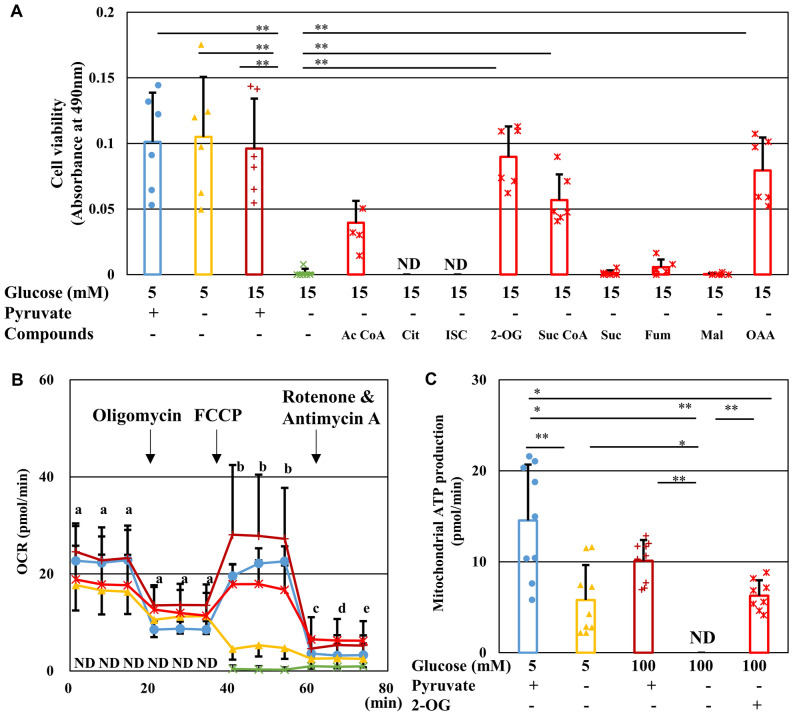
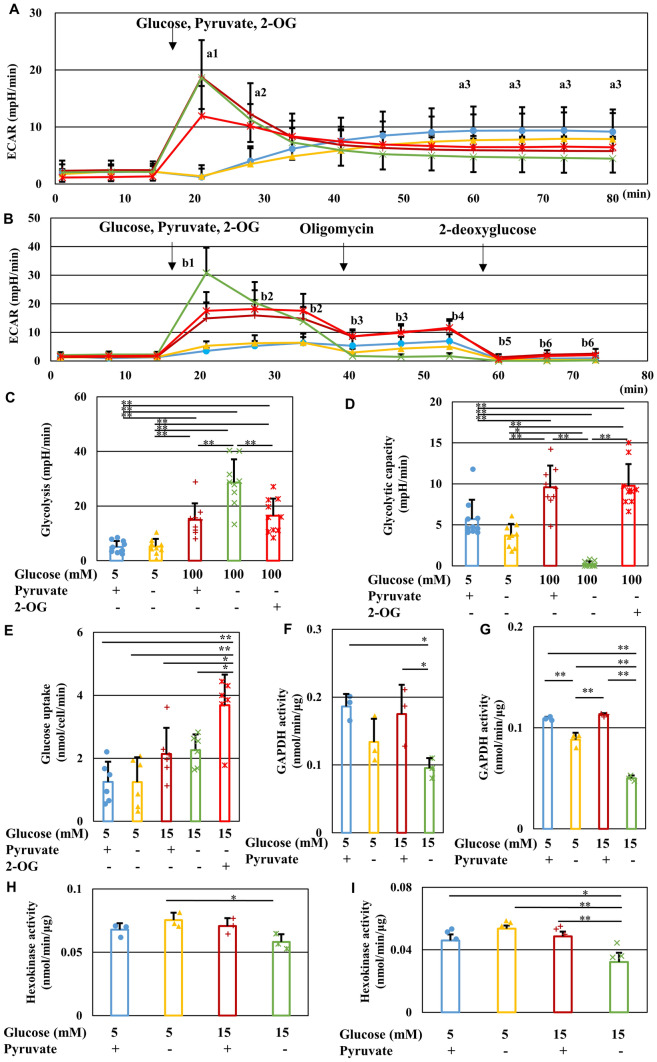
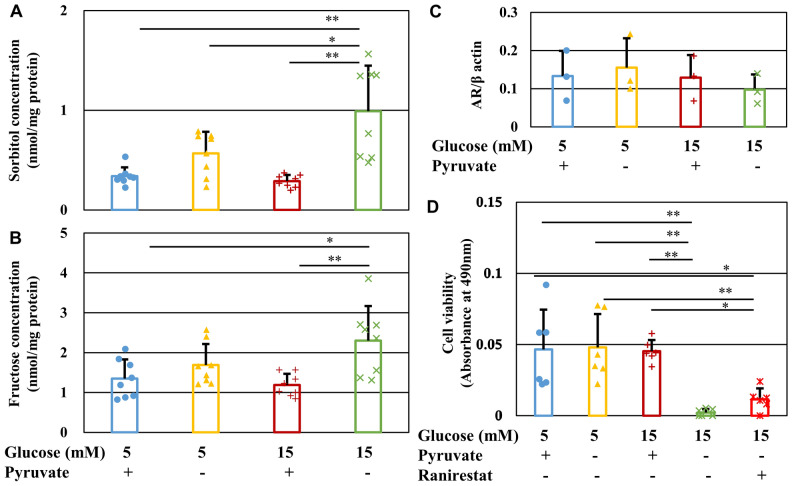
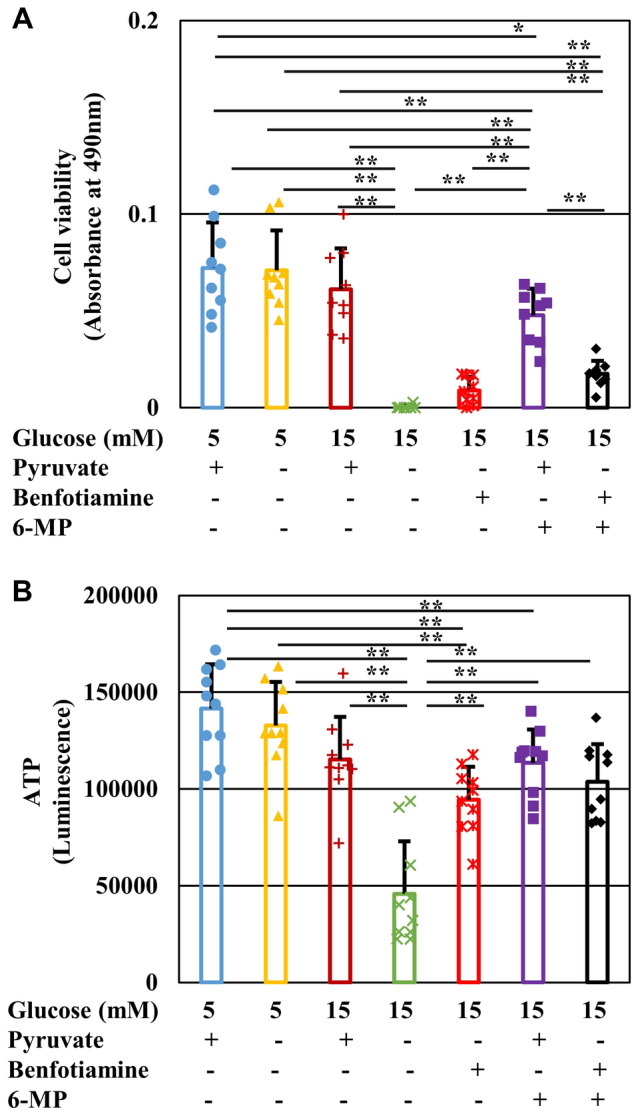
Pyruvate functions as a key molecule in energy production and as an antioxidant. The efficacy of pyruvate supplementation in diabetic retinopathy and nephropathy has been shown in animal models; however, its significance in the functional maintenance of neurons and Schwann cells under diabetic conditions remains unknown. We observed rapid and extensive cell death under high-glucose (> 10 mM) and pyruvate-starved conditions. Exposure of Schwann cells to these conditions led to a significant decrease in glycolytic flux, mitochondrial respiration and ATP production, accompanied by enhanced collateral glycolysis pathways (e.g., polyol pathway). Cell death could be prevented by supplementation with 2-oxoglutarate (a TCA cycle intermediate), benfotiamine (the vitamin B1 derivative that suppresses the collateral pathways), or the poly (ADP-ribose) polymerase (PARP) inhibitor, rucaparib. Our findings suggest that exogenous pyruvate plays a pivotal role in maintaining glycolysis-TCA cycle flux and ATP production under high-glucose conditions by suppressing PARP activity.
© 2021. The Author(s).
Conflict of interest statement
Research funding for this study, as well as the AR inhibitor ranirestat and the data of intracellular sorbitol and fructose contents, were provided by Sumitomo Dainippon Pharma Co., Ltd (H. Y. and K. S.). The other authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
